










Services on Demand
Journal
Article
 Spanish (pdf)
Spanish (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mailIndicators
-
 Cited by SciELO
Cited by SciELO -
 Access statistics
Access statistics
Related links
-
 Cited by Google
Cited by Google -
 Similars in
SciELO
Similars in
SciELO -
 Similars in Google
Similars in Google
Share

 Permalink
PermalinkRevista Colombiana de Ciencias Pecuarias
Print version ISSN 0120-0690On-line version ISSN 2256-2958
Rev Colom Cienc Pecua vol.20 no.2 Medellín Apr./June 2007
Modelación de la cinética de degradaciónde alimentos para rumiantes
Modeling of ruminant food degradation kinetics
Ricardo Rosero Noguera1*, Zoot Ph.D; Sandra L Posada Ochoa1, Zoot MS.
1Grupo de Investigación en Ciencias Agrarias, GRICA , Facultad de Ciencias Agrarias, Universidad de Antioquia, Medellín, Colombia
ricardonoguera@agronica.udea.edu.co
(Recibido: 31 mayo, 2006; aceptado: 24 mayo, 2007)
Resumen
La técnica de degradación in situ ha sido ampliamente adoptada para evaluar la tasa y la extensión de la degradación de los alimentos en el rumen. Los nuevos modelos para formulación y evaluación de raciones requieren de la determinación precisa de aspectos dinámicos de la degradación de alimentos en el rumen y han adoptado a la técnica in situ como instrumento para facilitar este tipo de mediciones. Numerosas son las fuentes de variación relacionadas con la técnica que pueden alterar las estimativas de la tasa y la extensión de la degradación, entre ellas podemos citar: el tamaño de los sacos de nylon, el tamaño de los poros del saco, la posición de los sacos dentro del rumen, el tamaño de partícula del alimento y el procesamiento matemático de la información. Diferentes modelos matemáticos han sido propuestos para estudiar la cinética de degradación. En la construcción de estos modelos han sido tenidas en cuenta las características particulares del sustrato en estudio, aspectos anatómicos y fisiológicos del animal y procesos relacionados con la hidratación y colonización del sustrato por parte de los microorganismos ruminales. El objetivo de este trabajo fue discutir algunos modelos utilizados en el estudio de la cinética de degradación del alimento mediante su descripción matemática y aplicación práctica.
Palabras clave: fermentación, producción de gas, rumiantes.
Summary
The in situ degradation technique has been adopted to evaluate the rate and extent of ruminal food degradation. The new models for formulation and evaluation of foods require of the precise determination of the dynamic aspects of ruminal foods degradation for which the in situ technique has been adopted in order to facilitate its mathematical determination. Different sources of variation related with the technique itself can alter the rate and extent of food degradation: the size of nylon bag, pore size, specific placement of the bag into the rumen, particle size of food and the mathematical models used for analyses, among others. Different models have been proposed for degradation kinetic studies. In these models the characteristics of food samples, anatomical and physiologic aspects of the animal, as well as those processes related with the hydrate and colonization of samples by ruminal microorganism have been considered. The objective of this work was to discuss some of the models used to evaluate the rate and extent of food degradation in the rumen by the mathematical description and practical application of the in situ technique.
Key words: fermentation, gas production, ruminants.
* Autor para el envío de correspondencia y solicitud de separatas: Grupo de Investigación en Ciencias Agrarias, Facultad de Ciencias Agrarias – GRICA Universidad de Antioquia, Medellín, Colombia. E-mail: ricardonoguera@agronica.udea.edu.co.
Introducción
La digestión de los rumiantes es un proceso complejo que involucra múltiples interacciones entre la dieta, los microorganismos ruminales y el hospedero. Separar el proceso en sus distintos componentes permite un mejor entendimiento de su dinámica y facilita su descripción matemática. Los modelos matemáticos permiten estudiar y estimar parámetros que describen la naturaleza intrínseca de los alimentos y las interacciones de los nutrientes que limitan su digestión.
Anatómicamente el tracto digestivo de los rumiantes puede ser dividido en tres compartimientos, cada uno con características propias y particulares: retículo-rumen, intestino delgado e intestino grueso. En el rumen y en el intestino grueso la digestión ocurre por acción microbiana, en tanto que, en el intestino delgado diferentes complejos enzimáticos degradan los componentes del alimento. De igual manera en el contenido ruminal pueden ser distinguidos dos subcompartimentos con diferentes características de degradación y pasaje: una fase líquida y una fase sólida en la que se evidencia la presencia de partículas con rápidas tasas de pasaje y degradación (alimentos concentrados) y partículas que presentan prolongados tiempos de retención y lenta degradación (forrajes).
Histológicamente los tejidos de las plantas forrajeras pueden ser divididos en tres tipos: 1) material rápidamente fermentable (células del mesófilo), 2) material de lenta fermentación (esclerénquima, parénquima) y 3) material indigestible (tejido vascular lignificado) (2). En las primeras horas de fermentación una parte del sustrato, principalmente los azúcares solubles son fermentados inmediatamente, sin embargo ellos sólo constituyen una pequeña parte del material potencialmente digestible. A medida que el proceso fermentativo continúa, una menor cantidad de material es hidratado y colonizado por los microorganismos ruminales lo que origina diferentes tasas de degradación dependiendo de la concentración de carbohidratos estructurales, contenido de lignina y estado de madurez de la planta.
Las tasas de digestión de los componentes de los alimentos varían considerablemente con valores que van desde 0.02 min-1 para azúcares solubles hasta 0.02 horas-1 para carbohidratos complejos en la fibra. Estas diferencias en las tasas de digestión y pasaje requieren que los componentes del alimento también sean identificados y contemplados en los modelos de digestión y degradación.
Los procesos de digestión y pasaje pueden ser descritos por modelos compartimentales en los cuales cada compartimiento representa un proceso distinto. Diferentes modelos han sido propuestos para describir la digestión y pasaje de los alimentos en los rumiantes (5, 6, 9, 12). En estos modelos, el alimento desaparece del rumen por degradación y absorción o por tránsito a tracto digestivo posterior apareciendo finalmente en las heces. La proporción de nutrientes que están disponibles para el rumiante varía en función de la competencia entre las tasas de degradación y pasaje.
La precisión de los modelos matemáticos para simular con buena aproximación situaciones reales, dependerá de contemplar en su construcción los factores anatómicos y fisiológicos del animal y los factores asociados al alimento como la composición química, palatabilidad y procesamiento. El objetivo de este trabajo fue discutir algunos modelos utilizados en el estudio de la cinética de degradación del alimento mediante su descripción matemática y aplicación práctica.
Métodos para estimar la tasa y la extensión de la degradación
Existen diferentes métodos que permiten estimar la tasa y la extensión de la degradación de los alimentos para predecir el valor nutricional de las forrajeras. Las técnicas in vitro permiten la evaluación rutinaria de la fermentación ruminal empleandofluidoruminalcomoenlatécnicadescrita por Tilley y Terry (23) o alternativamente sin la utilización de fluido ruminal sino con la utilización de complejos enzimáticos (4). Estos métodos ofrecen una estimativa de la digestibilidad potencial de los alimentos sin llevar en consideración los procesos de la dinámica ruminal. La digestibilidad in vivo históricamente ha sido utilizada para determinar la degradabilidad aparente de los alimentos, sin embargo, esta técnica no permite cuantificar qué fracción del alimento ha sido degradada en el rumen y qué fracción ha sido degradada en el sistema digestivo posterior.
Una segunda generación de métodos fue desarrollada incorporando las estimativas de la cinética de degradación en el retículo – rumen. Estas estimativas fueron realizadas a través de la técnica in situ (18, 24) o a través de la técnica de producción de gas (19). Estos métodos son ampliamente utilizados para evaluar el valor energético y proteico de los alimentos para rumiantes, su potencial de ingestión y la presencia de factores antinutricionales.
La técnica in situ ofrece la posibilidad de estudiar la degradabilidad ruminal de los alimentos a través de la utilización de sacos de nylon suspendidos en el rumen. Esta técnica ha sido adoptada por el AFRC (1) como método estándar para caracterizar la degradabilidad ruminal del nitrógeno. Esta técnica ha sido escogida debido a su gran aproximación a los resultados in vivo. Este método también puede ser usado para describir las características de degradación de los componentes estructurales del forraje.
Modelaje de la cinética de degradación
La cinética de degradación ruminal de la materia seca (MS), de nitrógeno (N) y de algunos constituyentes de la pared celular puede ser descrita a través de modelos no lineales. El modelo propuesto por Orskov y McDonald (18) para la degradación del N ha sido el más utilizado.

Donde:
P= degradabilidad potencial t = tiempo de incubación a = intercepto con el eje Y en el tiempo cero. Representa el sustrato soluble y completamente degradable que sale rápidamente del saco de nylon. b = la diferencia entre el intercepto (a) y la asíntota. Representa la fracción insoluble pero potencialmente degradable del sustrato el cual es degradado por los microorganismos de acuerdo con un proceso cinético de primer orden. c = tasa constante de la función b. 1- (a+b) = representa la fracción no degradable de la muestra.
La estimativa de los parámetros a, b y c se realiza a través de un proceso iterativo que busca la combinación de parámetros que produzca la menor suma de cuadrados del error (SQE). En otras palabras, la suma cuadrado total (SQT) en el análisis de varianza de la regresión no lineal es dividida en dos partes, una parte llamada suma de cuadrado de la regresión (o modelo) (SQM) y otra llamada suma de cuadrado del residuo (o error) (SQE). Como la SQT es siempre la misma para un conjunto de datos, el valor de la SQE depende exclusivamente del modelo, de tal forma que modelos que representan mejor a un conjunto de datos tendrán menores valores en la SQE.
La ecuación (i) supone la existencia de tres fracciones en el alimento. Una fracción no degradable que representa la fracción del alimento que permanece en el saco después de un prolongado tiempo de incubación, otra fracción insoluble pero potencialmente degradable por los microorganismos ruminales y una tercera fracción rápidamente degradable que además del material soluble incluye pequeñas partículas que pueden salir de los sacos de nylon.
Los parámetros estimados por la ecuación (i) pueden ser utilizados en conjunto con tasas de flujo para una dieta específica y estimar la degradabilidad efectiva de la muestra (ecuación ii) (18)

Donde: a, b y c fueron previamente definidos para la ecuación (i)
k = tasa de flujo de las partículas del rumen
Los aspectos clave para la estimación de la degradabiliad efectiva son el ajuste de los datos al modelo y la pertinencia de los horarios de incubación. Mertens (15) indica que los modelos para estudiar la cinética de degradación deben describir el proceso de digestión incluyendo tres fases durante el proceso, caracterizar cada una de estas fases 1) una fase lag o tiempo de colonización, 2) un dentro de la curva de degradación. Los periodo de rápida degradación y 3) una lenta intervalos óptimos y mínimos de incubación digestión proporcional al incremento de la fracción sugeridos por Mertens (15) para una adecuada indigestible. Los horarios de incubación de los sacos estimativa de los parámetros de degradación son de nylon en el rumen deben ir orientados a presentados en la tabla 1.

Durante el proceso de incubación existe un periodo donde ninguna o una reducida degradación del alimento ocurre, que es conocido como tiempo de colonización (lag phase). De acuerdo con Allen y Mertens (3) este tiempo de colonización es específico para cada alimento y representa el tiempo necesario para la hidratación del sustrato y la alteración física o química de la fibra que puede ser requerida antes de que las bacterias colonicen el sustrato y se inicie la actividad enzimática.
La correcta determinación del tiempo de colonización depende de la adecuada estimación de la fracción soluble de los alimentos y la cuantificación de la pérdida de pequeñas partículas que puedan escapar del saco de nylon en el tiempo cero. Weisbjerg et al (25) sugieren un posible método para corregir los datos de degradabilidad por la pérdida de pequeñas partículas en el tiempo cero:

Asumiendo que S + PQ = a
Donde: DPCti = degradabilidad potencial en el tiempo ti ti=tiempo inicial o tiempo cero a=previamente definida en la ecuación (i)
PQ=partículas pequeñas contenidas en la fracción “a” S= material soluble en la fracción “a”
Este método consiste en dividir la fracción a en dos subfracciones, una fracción soluble en agua y una segunda fracción que puede pasar a través de los poros del saco de nylon, se considera que esta última fracción se degrada a la misma tasa que las partículas que permanecen dentro de los sacos de nylon. El residuo que permanece en cada saco de nylon después de cada tiempo de incubación es corregido adicionando la cantidad de partículas digestibles que podrían salir de los sacos en un tiempo determinado. Huntington y Givens (10) consideran que este método para corregir los datos de degradabilidad para las partículas que consiguen atravesar los sacos de nylon puede ser útil para mejorar la exactitud de la técnica y la estimativa del tiempo de colonización, no obstante este tipo de corrección está basada en la hipótesis de que la perdida de partículas sea representativa lo que no siempre es verdadero.
La influencia del tiempo de colonización sobre la degradación de la proteína fue considerada por McDonald (12), quien sugirió el siguiente modelo para estimar el tiempo de colonización:
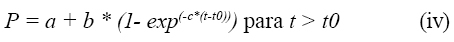
Donde: a, b y c fueron previamente definidos en la ecuación (i)
t = tiempo de incubación
ti= tiempo de colonización expresado en horas
El tiempo de colonización varía en función del sustrato degradado y la presencia de carbohidratos solubles. Noguera et al (16) incubando in vitro pasto kikuyo con diferentes niveles de almidón de papa encontraron que a medida que las concentraciones de carbohidratos solubles aumentan en la dieta, se reducen los tiempos de colonización y se incrementa la eficiencia en la síntesis de proteína microbiana.
Dhanoa (8) estableció que cuando el tiempo de colonización no es considerado en el modelo, existe una linearización de la curva de degradación que resulta en una subestimación de la tasa de degradación y una superestimación de la asíntota. Para superar este inconveniente este autor propone que la degradabilidad efectiva puede ser calculada a través de la siguiente expresión matemática:
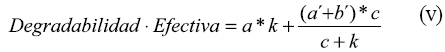
Donde: a, c y k fueron definidas previamente a´ y b´ son constantes.
Dhanoa et al (7) propusieron un modelo compartimental no linear para describir la cinética de degradación de las forrajeras, puesto que el modelo propuesto por Orskov y McDonald (18) fue desarrollado con datos obtenidos de la degradación de la proteína de la soja, que estaría completa después de 24 horas de incubación y podría ser estimada satisfactoriamente a través de ecuaciones de primer orden. Sin embargo, las forrajeras requieren periodos de incubación prolongados por lo que la utilización de este tipo de ecuaciones no sería el más apropiado, principalmente porque las tasas de degradación serían individuales para cada fracción, en especial con materiales altamente lignificados.
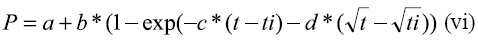
Donde: P= material degradado en el tiempo t a = fracción soluble b = fracción potencialmente degradable c y d = tasas de degradación de las fracciones a y b t = tiempo de incubación ti = tiempo de colonización expresado en horas.
En el caso que el parámetro d sea igual a cero el modelo propuesto por Dhanoa et al (7) sería igual a los modelos (i) y (iv).
En experimentos in vitro, Smith et al (22) determinaron la cinética de degradación de la pared celular de forrajeras asumiendo un tiempo de colonización igual a cero, mediante el siguiente modelo matemático:
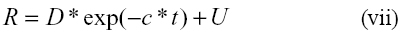
Donde: R= residuo en el tiempo t
D= fracción degradable
c= tasa constante de degradación
U= fracción indigestible
Posteriormente, Mertens y Loften (13) sugirieron la inclusión de un tiempo de colonización dentro del modelo para estudiar la cinética de degradación de la fibra in vitro:

Donde: R, D, c y U fueron definidas previamente en la ecuación (vii) L = tiempo de colonización
Cálculo de la degradabilidad efectiva
La técnica in situ ofrece la posibilidad de determinar la tasa y la extensión de la degradación de los alimentos. Sin embargo, la tasa de pasaje relativa a la tasa de digestión es una propiedad dinámica que afecta la digestibilidad.
En el rumen todas las partículas tienen la oportunidad de salir sin ser degradadas, la extensión de la degradación en los sacos de nylon no proporciona una adecuada estimativa de la degradación efectiva sobre condiciones normales. Por este motivo es necesario considerar conjuntamente la tasa de degradación y el flujo de las partículas dentro del rumen en la técnica in situ (17). Para estimar la degradabilidad efectiva de los alimentos diferentes métodos matemáticos han sido propuestos.
Kristensen et al (11) propone fijar una tasa de pasaje y sumar paso a paso la cantidad de alimento degradado durante cada periodo de incubación. En este método la tasa de degradación y la tasa de pasaje son combinadas para expresar la degradabilidad efectiva por la siguiente ecuación:
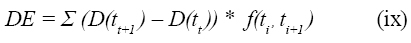
Donde, (D(tt+1 )–D(tt )) es la cantidad de alimento degradado durante el intervalo de tiempo ti a ti+1 , y f(ti’ ti+1 ) es la proporción de alimento que permanece em el rumen en el intervalo de tiempo ti a ti+1. La cantidad de alimento que permanece en el rumen (f(ti )) es estimada a partir da tasa de pasaje (k, h-1) por la siguiente ecuación:

La degradabilidad efectiva también puede ser calculada a partir de los parámetros obtenidos del ajuste exponencial de los datos, ecuaciones (i) y (iv) (8):
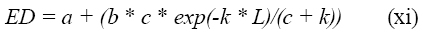
Donde: a, b, c y L fueron definidos en las ecuaciones (i) y (iv)
k corresponde a la tasa de pasaje (h-1).
De acuerdo con Noziére y Doreau (17) la proporcionalidad en el cálculo de la degradabilidad efectiva entre estos dos métodos depende del ajuste de los datos a la regresión y de la pertinencia de los horarios de incubación.
Ajuste de valores de degradación ruminal a un modelo de regresión no lineal
Dos métodos son utilizados para ajustar los datos a los modelos de la cinética ruminal: el primero, consiste en la transformación logarítmica de los datos, seguida por una regresión lineal; el segundo método, hace uso de la regresión no lineal. Para efectos demostrativos haremos uso del procedimiento PROC NLIN (nonlinear regression models) del paquete estadístico SAS (20). Los datos ajustados al modelo propuesto por Orskov y McDonald (18) (ecuación i) fueron tomados de un ejemplo numérico propuesto por Mertens (15).

A continuación se presenta la salida del paquete estadístico SAS después del ajuste al modelo no lineal de los datos propuestos:



Interpretación de la salida: puede observarse que el modelo alcanzo el criterio
1. Lista de suma de cuadrados residuales asociadas con todas o algunas de las combinaciones iniciales de los parámetros.
2. Estimación de los parámetros y las sumas de cuadrado del residuo determinadas en cada iteración.
3. Tabla de análisis de varianza incluyendo como fuentes de variación regresión, residual, total no corregido y total corregido.
4. Parámetros estimados.
5. Error estándar de los valores estimados.
6. Intervalo de confianza asintótico al 95% para los parámetros estimados.
7. Matriz de correlación para los parámetros estimados.
Después del ajuste de los datos al modelo propuesto por Orskov y McDonald (18) (ecuación (i)) de convergencia. El modelo estimó que la fracción soluble del sustrato (fracción a) fue de 0.76%, la fracción insoluble pero potencialmente degradable (fracción b) fue de 93.10%, la fracción indigestible (100- a + b) fue de 6.13%. La tasa de degradación de la fracción potencialmente degradable estimada fue de 9.65% h-1 y la degradabilidad potencial del sustrato después de 96 horas de incubación ruminal (a + b) fue de 93.86%.
Para estimar la extensión de la degradación del sustrato es necesario aplicar el modelo propuesto por Orskov y McDonald (18) (ecuación (ii)). Este modelo requiere considerar una tasa de pasaje para la fracción potencialmente degradable (b). Para nuestro ejemplo consideraremos una tasa de pasaje del 5% h-1.
Degradabilidad efectiva = a + ((b*c)/ (c + k))
Remplazando los valores de a, b y c estimados anteriormente obtenemos que la degradabilidad efectiva del sustrato con una tasa de pasaje del 5% h-1 fue de 61.40%.
Consideraciones finales
Los procesos de degradación y transito de las partículas por el tracto gastrointestinal son afectados por factores inherentes al animal y características intrínsecas del alimento. Estos factores deben ser considerados en los modelos matemáticos para estimar con mayor precisión los parámetros de la cinética de degradación del alimento en el tracto digestivo.
De igual manera, es importante resaltar que el ajuste de los modelos varía en función del ambiente de fermentación y la naturaleza del sustrato en estudio (forrajes vs. concentrados). De esta forma, en ensayos de degradabilidad in situ es necesaria la evaluación de diferentes modelos matemáticos con miras a mejorar la precisión de las predicciones.
La modelación de la cinética de degradación es solo una de las fuentes de variación que pueden afectar la tasa y la extensión de la degradación. En ensayos de digestibilidad in situ estos parámetros pueden ser directamente afectados por los horarios de incubación escogidos para construir la curva y el número de unidades experimentales utilizadas para este tipo de estudios.
El mayor limitante en la producción bovina de los países en desarrollo es la fluctuación en la cantidad y la calidad de los recursos forrajeros, por tanto, la descripción de los alimentos en términos de la cinética de digestión provee una base útil para su evaluación y eficiente utilización.
Referencias
1. AFRC. Agricultural and Food Research Council: Technical commite on responses to nutrients: Nutritive requeriments of ruminant animal protein. Nutr Abs Rev 1992; 68:65-71. [ Links ]
2. Akin DE. Microscopic evaluation of forage by rumen microorganisms – A review. J Anim Sci 1979; 48:701-722. [ Links ]
3. Allen MS, Mertens DR. Evaluating constraints on fiber digestion by rumen microbes. J Nutr 1988; 118:261-270. [ Links ]
4. Aufere J. Utilization des enzymes cellulolytiques pour prévior la digestibilité des forages. Bull Tech C R Z V 1982; 49:23-25. [ Links ]
5. Baldwin RL, Koong LJ, Ulyatt MJ. A dynamic model of ruminant digestion for evaluation of factors affecting nutritive value. Agric Syst 2 1977; 255-287. [ Links ]
6. Baldwin RL, Thornley JH, Beever DE. Lactating cow metabolism. 2. Digestive elements of a mechanistic model. J Dairy Res 1987; 54:107-131. [ Links ]
7. Dhanoa MS, France J, Siddons J, Lopez S, Buchanan-Smith JG. A non linear compartimental model to describe forage degradation kinetics during incubation in polyestes bags in the rumen. Br J Nutr 1995; 51:173-179. [ Links ]
8. Dhanoa MS. On the analysis of dacron bag data fir low degradability feeds. Grass Forage Sci 1988; 43:441-444. [ Links ]
9. France J, Thornley JH, Beever DE. A mathematical model of the rumen. J Agric Sci 1982; 99:343-353. [ Links ]
10. Huntington JA, Givens DI. Studies on In situ degradation of feeds in the rumen: 1- effect of species, bag motility and incubation squence on dry matter disappearance. An Feed Sci Techn 1997;64:227-241. [ Links ]
11. Kristensen ES, Moller PD, Hvelplund T. Estimation of the effective protein degradability in the rumen of cows using the nylon bag technique combined with the outflow rate. Acta Agr Scand 1982; 32:123-127[ [ Links ]STANDARDIZEDENDPARAG]
12. McDonald IM. A revised model for the estimation of protein degradability in rumen. J Agr Sci 1981; 96:251-252. [ Links ]
13. Mertens DR, Loften JR. The effect of starch on forage fiber digestion kinetics in vitro. J Dairy Sci 1980; 63:1437-1446. [ Links ]
14. Mertens DR, Ely LO. A dynamic model of fiber digestion and passage in the rumen for evaluating forage quality. J Anim Sci 1979; 49:1085-1095. [ Links ]
15. Mertens DR. Rate and extent of digestion. In: Forbes JM, France J, editors. Quantitative aspects of ruminant digestion and metabolism. UK, CAB International;1993. p. 13-51. [ Links ]
16. Noguera RR, Ramírez IC, Bolivar DM. Efecto de la inclusión de papa (Solanum tuberosum) en la cinética de fermentación in vitro del pasto kikuyo (Pennisetum clandestinum). Livestock Res Rural Develop 2006; Volume 18, Article # 64. Retrieved May 2, 2002, from http://www.cipav.org.co/lrrd/lrrd18/5/nogu18062.htm [ Links ]
Modeling of ruminant food degradation kinetics
Ricardo Rosero Noguera1*, Zoot Ph.D; Sandra L Posada Ochoa1, Zoot MS.
1Grupo de Investigación en Ciencias Agrarias, GRICA , Facultad de Ciencias Agrarias, Universidad de Antioquia, Medellín, Colombia
ricardonoguera@agronica.udea.edu.co
(Recibido: 31 mayo, 2006; aceptado: 24 mayo, 2007)
Resumen
La técnica de degradación in situ ha sido ampliamente adoptada para evaluar la tasa y la extensión de la degradación de los alimentos en el rumen. Los nuevos modelos para formulación y evaluación de raciones requieren de la determinación precisa de aspectos dinámicos de la degradación de alimentos en el rumen y han adoptado a la técnica in situ como instrumento para facilitar este tipo de mediciones. Numerosas son las fuentes de variación relacionadas con la técnica que pueden alterar las estimativas de la tasa y la extensión de la degradación, entre ellas podemos citar: el tamaño de los sacos de nylon, el tamaño de los poros del saco, la posición de los sacos dentro del rumen, el tamaño de partícula del alimento y el procesamiento matemático de la información. Diferentes modelos matemáticos han sido propuestos para estudiar la cinética de degradación. En la construcción de estos modelos han sido tenidas en cuenta las características particulares del sustrato en estudio, aspectos anatómicos y fisiológicos del animal y procesos relacionados con la hidratación y colonización del sustrato por parte de los microorganismos ruminales. El objetivo de este trabajo fue discutir algunos modelos utilizados en el estudio de la cinética de degradación del alimento mediante su descripción matemática y aplicación práctica.
Palabras clave: fermentación, producción de gas, rumiantes.
Summary
The in situ degradation technique has been adopted to evaluate the rate and extent of ruminal food degradation. The new models for formulation and evaluation of foods require of the precise determination of the dynamic aspects of ruminal foods degradation for which the in situ technique has been adopted in order to facilitate its mathematical determination. Different sources of variation related with the technique itself can alter the rate and extent of food degradation: the size of nylon bag, pore size, specific placement of the bag into the rumen, particle size of food and the mathematical models used for analyses, among others. Different models have been proposed for degradation kinetic studies. In these models the characteristics of food samples, anatomical and physiologic aspects of the animal, as well as those processes related with the hydrate and colonization of samples by ruminal microorganism have been considered. The objective of this work was to discuss some of the models used to evaluate the rate and extent of food degradation in the rumen by the mathematical description and practical application of the in situ technique.
Key words: fermentation, gas production, ruminants.
* Autor para el envío de correspondencia y solicitud de separatas: Grupo de Investigación en Ciencias Agrarias, Facultad de Ciencias Agrarias – GRICA Universidad de Antioquia, Medellín, Colombia. E-mail: ricardonoguera@agronica.udea.edu.co.
Introducción
La digestión de los rumiantes es un proceso complejo que involucra múltiples interacciones entre la dieta, los microorganismos ruminales y el hospedero. Separar el proceso en sus distintos componentes permite un mejor entendimiento de su dinámica y facilita su descripción matemática. Los modelos matemáticos permiten estudiar y estimar parámetros que describen la naturaleza intrínseca de los alimentos y las interacciones de los nutrientes que limitan su digestión.
Anatómicamente el tracto digestivo de los rumiantes puede ser dividido en tres compartimientos, cada uno con características propias y particulares: retículo-rumen, intestino delgado e intestino grueso. En el rumen y en el intestino grueso la digestión ocurre por acción microbiana, en tanto que, en el intestino delgado diferentes complejos enzimáticos degradan los componentes del alimento. De igual manera en el contenido ruminal pueden ser distinguidos dos subcompartimentos con diferentes características de degradación y pasaje: una fase líquida y una fase sólida en la que se evidencia la presencia de partículas con rápidas tasas de pasaje y degradación (alimentos concentrados) y partículas que presentan prolongados tiempos de retención y lenta degradación (forrajes).
Histológicamente los tejidos de las plantas forrajeras pueden ser divididos en tres tipos: 1) material rápidamente fermentable (células del mesófilo), 2) material de lenta fermentación (esclerénquima, parénquima) y 3) material indigestible (tejido vascular lignificado) (2). En las primeras horas de fermentación una parte del sustrato, principalmente los azúcares solubles son fermentados inmediatamente, sin embargo ellos sólo constituyen una pequeña parte del material potencialmente digestible. A medida que el proceso fermentativo continúa, una menor cantidad de material es hidratado y colonizado por los microorganismos ruminales lo que origina diferentes tasas de degradación dependiendo de la concentración de carbohidratos estructurales, contenido de lignina y estado de madurez de la planta.
Las tasas de digestión de los componentes de los alimentos varían considerablemente con valores que van desde 0.02 min-1 para azúcares solubles hasta 0.02 horas-1 para carbohidratos complejos en la fibra. Estas diferencias en las tasas de digestión y pasaje requieren que los componentes del alimento también sean identificados y contemplados en los modelos de digestión y degradación.
Los procesos de digestión y pasaje pueden ser descritos por modelos compartimentales en los cuales cada compartimiento representa un proceso distinto. Diferentes modelos han sido propuestos para describir la digestión y pasaje de los alimentos en los rumiantes (5, 6, 9, 12). En estos modelos, el alimento desaparece del rumen por degradación y absorción o por tránsito a tracto digestivo posterior apareciendo finalmente en las heces. La proporción de nutrientes que están disponibles para el rumiante varía en función de la competencia entre las tasas de degradación y pasaje.
La precisión de los modelos matemáticos para simular con buena aproximación situaciones reales, dependerá de contemplar en su construcción los factores anatómicos y fisiológicos del animal y los factores asociados al alimento como la composición química, palatabilidad y procesamiento. El objetivo de este trabajo fue discutir algunos modelos utilizados en el estudio de la cinética de degradación del alimento mediante su descripción matemática y aplicación práctica.
Métodos para estimar la tasa y la extensión de la degradación
Existen diferentes métodos que permiten estimar la tasa y la extensión de la degradación de los alimentos para predecir el valor nutricional de las forrajeras. Las técnicas in vitro permiten la evaluación rutinaria de la fermentación ruminal empleandofluidoruminalcomoenlatécnicadescrita por Tilley y Terry (23) o alternativamente sin la utilización de fluido ruminal sino con la utilización de complejos enzimáticos (4). Estos métodos ofrecen una estimativa de la digestibilidad potencial de los alimentos sin llevar en consideración los procesos de la dinámica ruminal. La digestibilidad in vivo históricamente ha sido utilizada para determinar la degradabilidad aparente de los alimentos, sin embargo, esta técnica no permite cuantificar qué fracción del alimento ha sido degradada en el rumen y qué fracción ha sido degradada en el sistema digestivo posterior.
Una segunda generación de métodos fue desarrollada incorporando las estimativas de la cinética de degradación en el retículo – rumen. Estas estimativas fueron realizadas a través de la técnica in situ (18, 24) o a través de la técnica de producción de gas (19). Estos métodos son ampliamente utilizados para evaluar el valor energético y proteico de los alimentos para rumiantes, su potencial de ingestión y la presencia de factores antinutricionales.
La técnica in situ ofrece la posibilidad de estudiar la degradabilidad ruminal de los alimentos a través de la utilización de sacos de nylon suspendidos en el rumen. Esta técnica ha sido adoptada por el AFRC (1) como método estándar para caracterizar la degradabilidad ruminal del nitrógeno. Esta técnica ha sido escogida debido a su gran aproximación a los resultados in vivo. Este método también puede ser usado para describir las características de degradación de los componentes estructurales del forraje.
Modelaje de la cinética de degradación
La cinética de degradación ruminal de la materia seca (MS), de nitrógeno (N) y de algunos constituyentes de la pared celular puede ser descrita a través de modelos no lineales. El modelo propuesto por Orskov y McDonald (18) para la degradación del N ha sido el más utilizado.
Donde:
P= degradabilidad potencial t = tiempo de incubación a = intercepto con el eje Y en el tiempo cero. Representa el sustrato soluble y completamente degradable que sale rápidamente del saco de nylon. b = la diferencia entre el intercepto (a) y la asíntota. Representa la fracción insoluble pero potencialmente degradable del sustrato el cual es degradado por los microorganismos de acuerdo con un proceso cinético de primer orden. c = tasa constante de la función b. 1- (a+b) = representa la fracción no degradable de la muestra.
La estimativa de los parámetros a, b y c se realiza a través de un proceso iterativo que busca la combinación de parámetros que produzca la menor suma de cuadrados del error (SQE). En otras palabras, la suma cuadrado total (SQT) en el análisis de varianza de la regresión no lineal es dividida en dos partes, una parte llamada suma de cuadrado de la regresión (o modelo) (SQM) y otra llamada suma de cuadrado del residuo (o error) (SQE). Como la SQT es siempre la misma para un conjunto de datos, el valor de la SQE depende exclusivamente del modelo, de tal forma que modelos que representan mejor a un conjunto de datos tendrán menores valores en la SQE.
La ecuación (i) supone la existencia de tres fracciones en el alimento. Una fracción no degradable que representa la fracción del alimento que permanece en el saco después de un prolongado tiempo de incubación, otra fracción insoluble pero potencialmente degradable por los microorganismos ruminales y una tercera fracción rápidamente degradable que además del material soluble incluye pequeñas partículas que pueden salir de los sacos de nylon.
Los parámetros estimados por la ecuación (i) pueden ser utilizados en conjunto con tasas de flujo para una dieta específica y estimar la degradabilidad efectiva de la muestra (ecuación ii) (18)
Donde: a, b y c fueron previamente definidos para la ecuación (i)
k = tasa de flujo de las partículas del rumen
Los aspectos clave para la estimación de la degradabiliad efectiva son el ajuste de los datos al modelo y la pertinencia de los horarios de incubación. Mertens (15) indica que los modelos para estudiar la cinética de degradación deben describir el proceso de digestión incluyendo tres fases durante el proceso, caracterizar cada una de estas fases 1) una fase lag o tiempo de colonización, 2) un dentro de la curva de degradación. Los periodo de rápida degradación y 3) una lenta intervalos óptimos y mínimos de incubación digestión proporcional al incremento de la fracción sugeridos por Mertens (15) para una adecuada indigestible. Los horarios de incubación de los sacos estimativa de los parámetros de degradación son de nylon en el rumen deben ir orientados a presentados en la tabla 1.
Durante el proceso de incubación existe un periodo donde ninguna o una reducida degradación del alimento ocurre, que es conocido como tiempo de colonización (lag phase). De acuerdo con Allen y Mertens (3) este tiempo de colonización es específico para cada alimento y representa el tiempo necesario para la hidratación del sustrato y la alteración física o química de la fibra que puede ser requerida antes de que las bacterias colonicen el sustrato y se inicie la actividad enzimática.
La correcta determinación del tiempo de colonización depende de la adecuada estimación de la fracción soluble de los alimentos y la cuantificación de la pérdida de pequeñas partículas que puedan escapar del saco de nylon en el tiempo cero. Weisbjerg et al (25) sugieren un posible método para corregir los datos de degradabilidad por la pérdida de pequeñas partículas en el tiempo cero:
Asumiendo que S + PQ = a
Donde: DPCti = degradabilidad potencial en el tiempo ti ti=tiempo inicial o tiempo cero a=previamente definida en la ecuación (i)
PQ=partículas pequeñas contenidas en la fracción “a” S= material soluble en la fracción “a”
Este método consiste en dividir la fracción a en dos subfracciones, una fracción soluble en agua y una segunda fracción que puede pasar a través de los poros del saco de nylon, se considera que esta última fracción se degrada a la misma tasa que las partículas que permanecen dentro de los sacos de nylon. El residuo que permanece en cada saco de nylon después de cada tiempo de incubación es corregido adicionando la cantidad de partículas digestibles que podrían salir de los sacos en un tiempo determinado. Huntington y Givens (10) consideran que este método para corregir los datos de degradabilidad para las partículas que consiguen atravesar los sacos de nylon puede ser útil para mejorar la exactitud de la técnica y la estimativa del tiempo de colonización, no obstante este tipo de corrección está basada en la hipótesis de que la perdida de partículas sea representativa lo que no siempre es verdadero.
La influencia del tiempo de colonización sobre la degradación de la proteína fue considerada por McDonald (12), quien sugirió el siguiente modelo para estimar el tiempo de colonización:
Donde: a, b y c fueron previamente definidos en la ecuación (i)
t = tiempo de incubación
ti= tiempo de colonización expresado en horas
El tiempo de colonización varía en función del sustrato degradado y la presencia de carbohidratos solubles. Noguera et al (16) incubando in vitro pasto kikuyo con diferentes niveles de almidón de papa encontraron que a medida que las concentraciones de carbohidratos solubles aumentan en la dieta, se reducen los tiempos de colonización y se incrementa la eficiencia en la síntesis de proteína microbiana.
Dhanoa (8) estableció que cuando el tiempo de colonización no es considerado en el modelo, existe una linearización de la curva de degradación que resulta en una subestimación de la tasa de degradación y una superestimación de la asíntota. Para superar este inconveniente este autor propone que la degradabilidad efectiva puede ser calculada a través de la siguiente expresión matemática:
Donde: a, c y k fueron definidas previamente a´ y b´ son constantes.
Dhanoa et al (7) propusieron un modelo compartimental no linear para describir la cinética de degradación de las forrajeras, puesto que el modelo propuesto por Orskov y McDonald (18) fue desarrollado con datos obtenidos de la degradación de la proteína de la soja, que estaría completa después de 24 horas de incubación y podría ser estimada satisfactoriamente a través de ecuaciones de primer orden. Sin embargo, las forrajeras requieren periodos de incubación prolongados por lo que la utilización de este tipo de ecuaciones no sería el más apropiado, principalmente porque las tasas de degradación serían individuales para cada fracción, en especial con materiales altamente lignificados.
Donde: P= material degradado en el tiempo t a = fracción soluble b = fracción potencialmente degradable c y d = tasas de degradación de las fracciones a y b t = tiempo de incubación ti = tiempo de colonización expresado en horas.
En el caso que el parámetro d sea igual a cero el modelo propuesto por Dhanoa et al (7) sería igual a los modelos (i) y (iv).
En experimentos in vitro, Smith et al (22) determinaron la cinética de degradación de la pared celular de forrajeras asumiendo un tiempo de colonización igual a cero, mediante el siguiente modelo matemático:
Donde: R= residuo en el tiempo t
D= fracción degradable
c= tasa constante de degradación
U= fracción indigestible
Posteriormente, Mertens y Loften (13) sugirieron la inclusión de un tiempo de colonización dentro del modelo para estudiar la cinética de degradación de la fibra in vitro:
Donde: R, D, c y U fueron definidas previamente en la ecuación (vii) L = tiempo de colonización
Cálculo de la degradabilidad efectiva
La técnica in situ ofrece la posibilidad de determinar la tasa y la extensión de la degradación de los alimentos. Sin embargo, la tasa de pasaje relativa a la tasa de digestión es una propiedad dinámica que afecta la digestibilidad.
En el rumen todas las partículas tienen la oportunidad de salir sin ser degradadas, la extensión de la degradación en los sacos de nylon no proporciona una adecuada estimativa de la degradación efectiva sobre condiciones normales. Por este motivo es necesario considerar conjuntamente la tasa de degradación y el flujo de las partículas dentro del rumen en la técnica in situ (17). Para estimar la degradabilidad efectiva de los alimentos diferentes métodos matemáticos han sido propuestos.
Kristensen et al (11) propone fijar una tasa de pasaje y sumar paso a paso la cantidad de alimento degradado durante cada periodo de incubación. En este método la tasa de degradación y la tasa de pasaje son combinadas para expresar la degradabilidad efectiva por la siguiente ecuación:
Donde, (D(tt+1 )–D(tt )) es la cantidad de alimento degradado durante el intervalo de tiempo ti a ti+1 , y f(ti’ ti+1 ) es la proporción de alimento que permanece em el rumen en el intervalo de tiempo ti a ti+1. La cantidad de alimento que permanece en el rumen (f(ti )) es estimada a partir da tasa de pasaje (k, h-1) por la siguiente ecuación:
La degradabilidad efectiva también puede ser calculada a partir de los parámetros obtenidos del ajuste exponencial de los datos, ecuaciones (i) y (iv) (8):
Donde: a, b, c y L fueron definidos en las ecuaciones (i) y (iv)
k corresponde a la tasa de pasaje (h-1).
De acuerdo con Noziére y Doreau (17) la proporcionalidad en el cálculo de la degradabilidad efectiva entre estos dos métodos depende del ajuste de los datos a la regresión y de la pertinencia de los horarios de incubación.
Ajuste de valores de degradación ruminal a un modelo de regresión no lineal
Dos métodos son utilizados para ajustar los datos a los modelos de la cinética ruminal: el primero, consiste en la transformación logarítmica de los datos, seguida por una regresión lineal; el segundo método, hace uso de la regresión no lineal. Para efectos demostrativos haremos uso del procedimiento PROC NLIN (nonlinear regression models) del paquete estadístico SAS (20). Los datos ajustados al modelo propuesto por Orskov y McDonald (18) (ecuación i) fueron tomados de un ejemplo numérico propuesto por Mertens (15).
A continuación se presenta la salida del paquete estadístico SAS después del ajuste al modelo no lineal de los datos propuestos:
Interpretación de la salida: puede observarse que el modelo alcanzo el criterio
1. Lista de suma de cuadrados residuales asociadas con todas o algunas de las combinaciones iniciales de los parámetros.
2. Estimación de los parámetros y las sumas de cuadrado del residuo determinadas en cada iteración.
3. Tabla de análisis de varianza incluyendo como fuentes de variación regresión, residual, total no corregido y total corregido.
4. Parámetros estimados.
5. Error estándar de los valores estimados.
6. Intervalo de confianza asintótico al 95% para los parámetros estimados.
7. Matriz de correlación para los parámetros estimados.
Después del ajuste de los datos al modelo propuesto por Orskov y McDonald (18) (ecuación (i)) de convergencia. El modelo estimó que la fracción soluble del sustrato (fracción a) fue de 0.76%, la fracción insoluble pero potencialmente degradable (fracción b) fue de 93.10%, la fracción indigestible (100- a + b) fue de 6.13%. La tasa de degradación de la fracción potencialmente degradable estimada fue de 9.65% h-1 y la degradabilidad potencial del sustrato después de 96 horas de incubación ruminal (a + b) fue de 93.86%.
Para estimar la extensión de la degradación del sustrato es necesario aplicar el modelo propuesto por Orskov y McDonald (18) (ecuación (ii)). Este modelo requiere considerar una tasa de pasaje para la fracción potencialmente degradable (b). Para nuestro ejemplo consideraremos una tasa de pasaje del 5% h-1.
Degradabilidad efectiva = a + ((b*c)/ (c + k))
Remplazando los valores de a, b y c estimados anteriormente obtenemos que la degradabilidad efectiva del sustrato con una tasa de pasaje del 5% h-1 fue de 61.40%.
Consideraciones finales
Los procesos de degradación y transito de las partículas por el tracto gastrointestinal son afectados por factores inherentes al animal y características intrínsecas del alimento. Estos factores deben ser considerados en los modelos matemáticos para estimar con mayor precisión los parámetros de la cinética de degradación del alimento en el tracto digestivo.
De igual manera, es importante resaltar que el ajuste de los modelos varía en función del ambiente de fermentación y la naturaleza del sustrato en estudio (forrajes vs. concentrados). De esta forma, en ensayos de degradabilidad in situ es necesaria la evaluación de diferentes modelos matemáticos con miras a mejorar la precisión de las predicciones.
La modelación de la cinética de degradación es solo una de las fuentes de variación que pueden afectar la tasa y la extensión de la degradación. En ensayos de digestibilidad in situ estos parámetros pueden ser directamente afectados por los horarios de incubación escogidos para construir la curva y el número de unidades experimentales utilizadas para este tipo de estudios.
El mayor limitante en la producción bovina de los países en desarrollo es la fluctuación en la cantidad y la calidad de los recursos forrajeros, por tanto, la descripción de los alimentos en términos de la cinética de digestión provee una base útil para su evaluación y eficiente utilización.
Referencias
1. AFRC. Agricultural and Food Research Council: Technical commite on responses to nutrients: Nutritive requeriments of ruminant animal protein. Nutr Abs Rev 1992; 68:65-71. [ Links ]
2. Akin DE. Microscopic evaluation of forage by rumen microorganisms – A review. J Anim Sci 1979; 48:701-722. [ Links ]
3. Allen MS, Mertens DR. Evaluating constraints on fiber digestion by rumen microbes. J Nutr 1988; 118:261-270. [ Links ]
4. Aufere J. Utilization des enzymes cellulolytiques pour prévior la digestibilité des forages. Bull Tech C R Z V 1982; 49:23-25. [ Links ]
5. Baldwin RL, Koong LJ, Ulyatt MJ. A dynamic model of ruminant digestion for evaluation of factors affecting nutritive value. Agric Syst 2 1977; 255-287. [ Links ]
6. Baldwin RL, Thornley JH, Beever DE. Lactating cow metabolism. 2. Digestive elements of a mechanistic model. J Dairy Res 1987; 54:107-131. [ Links ]
7. Dhanoa MS, France J, Siddons J, Lopez S, Buchanan-Smith JG. A non linear compartimental model to describe forage degradation kinetics during incubation in polyestes bags in the rumen. Br J Nutr 1995; 51:173-179. [ Links ]
8. Dhanoa MS. On the analysis of dacron bag data fir low degradability feeds. Grass Forage Sci 1988; 43:441-444. [ Links ]
9. France J, Thornley JH, Beever DE. A mathematical model of the rumen. J Agric Sci 1982; 99:343-353. [ Links ]
10. Huntington JA, Givens DI. Studies on In situ degradation of feeds in the rumen: 1- effect of species, bag motility and incubation squence on dry matter disappearance. An Feed Sci Techn 1997;64:227-241. [ Links ]
11. Kristensen ES, Moller PD, Hvelplund T. Estimation of the effective protein degradability in the rumen of cows using the nylon bag technique combined with the outflow rate. Acta Agr Scand 1982; 32:123-127[ [ Links ]STANDARDIZEDENDPARAG]
12. McDonald IM. A revised model for the estimation of protein degradability in rumen. J Agr Sci 1981; 96:251-252. [ Links ]
13. Mertens DR, Loften JR. The effect of starch on forage fiber digestion kinetics in vitro. J Dairy Sci 1980; 63:1437-1446. [ Links ]
14. Mertens DR, Ely LO. A dynamic model of fiber digestion and passage in the rumen for evaluating forage quality. J Anim Sci 1979; 49:1085-1095. [ Links ]
15. Mertens DR. Rate and extent of digestion. In: Forbes JM, France J, editors. Quantitative aspects of ruminant digestion and metabolism. UK, CAB International;1993. p. 13-51. [ Links ]
16. Noguera RR, Ramírez IC, Bolivar DM. Efecto de la inclusión de papa (Solanum tuberosum) en la cinética de fermentación in vitro del pasto kikuyo (Pennisetum clandestinum). Livestock Res Rural Develop 2006; Volume 18, Article # 64. Retrieved May 2, 2002, from http://www.cipav.org.co/lrrd/lrrd18/5/nogu18062.htm [ Links ]

