










Services on Demand
Journal
Article
 Spanish (pdf)
Spanish (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mailIndicators
-
 Cited by SciELO
Cited by SciELO -
 Access statistics
Access statistics
Related links
-
 Cited by Google
Cited by Google -
 Similars in
SciELO
Similars in
SciELO -
 Similars in Google
Similars in Google
Share

 Permalink
PermalinkRevista Colombiana de Ciencias Pecuarias
Print version ISSN 0120-0690On-line version ISSN 2256-2958
Rev Colom Cienc Pecua vol.24 no.2 Medellín Apr./June 2011
Adaptation to a commercial diet alters post-larvae growth and development of striped catfish (Pseudoplatystoma sp) and yaque (Leiarius marmoratus)¤
Crecimiento y sobrevivencia de post-larvas de bagre rayado (Pseudoplatystoma sp) y yaque (Leiarius marmoratus) consumiendo una dieta seca
Crescimento e sobrevivência de pós-larvas do pintado (Pseudoplatystoma sp) e yaque (Leiarius marmoratus) alimentadas com ração seca
Lili J Marciales-Caro1 Biol; Nubia E Cruz-Casallas1 Ing Agr, Esp; John J Díaz-Olarte1 Biol; Víctor M Medina-Robles1 MVZ, MSc; Pablo E Cruz-Casallas1* MVZ, MSc, PhD.
1Grupo de Investigación sobre Reproducción y Toxicología de Organismos Acuáticos (GRITOX), Instituto de Acuicultura, Universidad de los Llanos, A.A. 110, Villavicencio, Colombia.
(Recibido: 14 diciembre, 2009; aceptado: 26 abril, 2011)
¤ Para citar este artículo: Marciales-Caro LJ, Cruz-Casallas NE, Díaz-Olarte JJ, Medina-Robles VM, Cruz-Casallas PE. Crecimiento y sobrevivencia de post-larvas de bagre rayado (Pseudoplatystoma sp) y yaque (Leiarius marmoratus) consumiendo una dieta seca. .
* Autor para correspondencia: Pablo Emilio Cruz-Casallas. Universidad de los Llanos, Km 12 Vía Puerto López, Vereda Barcelona, Villavicencio, Colombia. E-mail: pecruzcasallas@unillanos.edu.co.
Summary
The main challenges facing fish farming include the optimization of Silurid fingerling production through efficient larval rearing and nursery management. This study evaluated growth performance of catfish (Pseudoplatystoma sp) and yaque (Leiarius marmoratus) post-larvae during adaptation to commercial diet containing 40% crude protein of animal origin (e.g., bovine liver and heart). A total of 240 L. marmoratus and Pseudoplatystoma sp. post-larvae were selected at 16 days of age. Postlarvae average weight and lengh were 57.2 ± 4.8 mg and 17.1 ± 1.4 mm for L. marmoratus, and 55.9 ± 6.6 mg and 13.5 ± 2.7 mm for Pseudoplatystoma sp, respectively. Housing density was 4 larvae / L. Four treatments (T) were evaluated, with five replicates each, as follows: T1: commercial feed (C), T2:C + bovine heart (CO), T3: C + liver (HI), and T4: C + liver + fish oil (HI + AP). The best weight gain for larvae of L. marmoratus was observed in T4 (3213 ± 278 mg). For Pseudoplatystoma sp., no significant differences were observed between treatments, although T2 was numerically better (695 ± 149 mg). In conclusion, conditioning to a dry diet was more effective for L. marmoratus post-larvae compared to Pseudoplatystoma sp. This is mainly due to the attractiveness and digestibility of the feed, as well as certain post-larvae characteristics (e.g., development of the digestive tract). It is necessary to conduct specific studies related to the ontogenetic development of the digestive tract to uncover the nutritional needs of each species during growth to match allowances with specific dietary requirements.
Key words: post-larvae, silurids, weaning.
Resumen
El principal desafío de la piscicultura con silúridos consiste en optimizar la producción de alevinos mediante prácticas eficientes de larvicultura y alevinaje. El presente estudio evaluó el desempeño productivo de post-larvas de bagre rayado (Pseudoplatystoma sp) y de yaque (Leiarius marmoratus) durante su acostumbramiento a una dieta comercial del 40% de proteína bruta de origen animal (ej: hígado y corazón de bovino). Fueron seleccionadas 240 post-larvas de L. marmoratus y de Pseudoplatystoma sp de 16 días de edad, con peso y talla promedio de 57.2 ± 4.8 mg y 17.1 ± 1.4 mm para L. marmoratus, y de 55.9 ± 6.6 mg y 13.5 ± 2.7 mm para Pseudoplatystoma sp, respectivamente. La densidad de alojamiento fue de 4 larvas/L. Fueron evaluados 4 tratamientos (T) con cinco réplicas cada uno, así: T1: alimento balanceado comercial (C); T2: C + corazón de bovino (CO); T3: C + hígado (HI) y; T4: C + hígado + aceite de pescado (HI+AP). La mejor ganancia de peso para larvas de L. marmoratus se observó en el T4 (3213 ± 278 mg). Para Pseudoplatystoma sp no se observaron diferencias significativas entre tratamientos, aunque el mejor resultado numérico fue para el T2 (695 ± 149 mg). En conclusión, el acondicionamiento a dieta seca fue más eficiente en post-larvas de L. marmoratus que en Pseudoplatystoma sp, debido principalmente a la atractabilidad y digestibilidad del alimento, así como a características de las post-larvas (desarrollo del tracto digestivo). Se hace necesario realizar estudios específicos de desarrollo ontogénico relacionados con las características del tracto digestivo que permitan evidenciar las necesidades nutricionales de cada especie durante su crecimiento, para formular dietas acordes con los requerimientos específicos.
Palabras Clave: acondicionamiento, post-larvas, silúridos.
Resumo
O principal desafio da aquicultura com silúridas é optimizar a produção de alevines mediante práticas eficientes de larvicultura e alevinagem. O presente estudo avaliou o desempenho produtivo de pós-larvas de pintado (Pseudoplatystoma sp) e Yaque (Leiarius marmoratus) durante o período de adaptação a uma dieta comercial com 40% de proteína bruta de origem animal (por exemplo, fígado e coração de bovino). Foram seleccionados 240 pós-larvas de L. marmoratus e Pseudoplatystoma sp 16 dias de idade, peso e comprimento médio de 57.2 ± 4.8 mg e 17.1 ± 1.4 mm para L. marmoratus, e 55.9 ± 6.6 mg e 13.5 ± 2.7 mm para Pseudoplatystoma sp, respectivamente. A densidade de alojamento foi de 4 larvas/L. Foram avaliados quatro tratamentos (T), com cinco repetições, conforme segue: T1: ração comercial (C), T2: C + coração (CO), T3: C + fígado (HI) e T4: C + fígado + óleo de peixe (HI + AP). O melhor ganho de peso para as larvas de L. marmoratus foi observado no T4 (3213 mg ± 278). Para Pseudoplatystoma sp não houve diferenças significativas entre os tratamentos, embora o resultado numérico foi melhor para o T2 (695 mg ± 149). Em conclusão, condicionado à dieta seca foi mais eficaz no pós-larvas de L. marmoratus que no Pseudoplatystoma sp, principalmente devido à atractividade e digestibilidade de alimentos para animais,bem como características de pós-larvas (desenvolvimento do trato digestivo). É necessário realizar estudos específicos relacionados com o desenvolvimento ontogênico das características do aparelho digestivo que revelam as necessidades nutricionais de cada espécie durante o seu crescimento para fazer concessões em conformidade com os requisitos específicos.
Palavras-chave: acondicionamento, pós-larvas, silúridas.
Introducción
En los peces, durante el desarrollo larvario se presentan cambios importantes en el sistema digestivo, involucrando procesos de digestión, absorción, transporte y asimilación de compuestos químicos y por lo tanto, determinando modificaciones en los requerimientos nutricionales de acuerdo con la etapa de desarrollo de la larva (Kolkovski, 2001). Con base en el grado de desarrollo del tracto digestivo, Dabrowski (1984) clasificó las larvas de los peces en tres tipos: 1) larvas con sistema digestivo ampliamente desarrollado antes de iniciar la alimentación endógena (salmónidos), 2) larvas con sistema digestivo con desarrollo intermedio de zonas de digestión y absorción, al momento de agotar las reservas del saco vitelino (cíclidos) y, 3) larvas con desarrollo tardío del estómago funcional, glándulas gástricas y demás órganos digestivos, tal como ocurre en la mayoría de las larvas de peces, incluyendo las de silúridos.
Debido a la ausencia de enzimas digestivas, este último grupo requiere incluir en su dieta alimento vivo, que le proporcione estímulos visuales y aportes enzimáticos, para incrementar la actividad digestiva e ingestiva (Aristizábal y Suárez, 2006; Kolkovsky, 2001) y permitir una más fácil transición a una dieta inerte (Borges y Portella, 2006).
Con relación a la piscicultura de silúridos, las prácticas piscícolas en estas especies tienen actualmente como desafío principal la optimización de la producción de alevinos, mejorando la eficiencia de los procesos de larvicultura y el alevinaje (Segura et al., 2004; Díaz-Olarte et al., 2009), siendo factores decisivos para este propósito el manejo adecuado de las fuentes de alimento, especialmente durante las primeras etapas de desarrollo, así como la implementación de dietas artificiales que se conviertan en la principal fuente de alimento. El proceso de acondicionamiento a consumo de alimentos artificiales, conocido comúnmente como destete, consiste en proporcionar una dieta mixta que contenga alimento artificial (alimento balanceado comercial) acompañado de un suplemento proteico de origen animal, el cual va disminuyéndose gradualmente hasta proporcionar solamente el alimento artificial o preparado comercial.
La implementación de este proceso proporciona ventajas desde el punto de vista económico, ya que disminuye el uso de organismos vivos como fuente de alimento (artemia salina o larvas de peces) y optimiza el uso de alimento balanceados comerciales (Guerrero-Alvarado, 2003). Este proceso ha sido logrado satisfactoriamente en Colossoma macropomum (Sevilla y Günther, 2000), Pangasius bocourti (Hung et al., 2002), Heterobranchus longifilis (Kerdchuen y Legendre, 1994), Sorubim cuspicaudus (Vergara y Hoyos, 2005) y Pseudoplatystoma corruscans (Guerrero-Alvarado, 2003); sin embargo, en especies nativas de la Orinoquia Colombiana aun no han sido publicados trabajos que describan este proceso, el cual es necesario para optimizar la larvicultura y el alevinaje de individuos de alto valor comercial como el bagre rayado y el yaque. Por lo tanto, el objetivo del presente trabajo fue evaluar el desempeño productivo de post-larvas de bagre rayado (Pseudoplatystoma sp) y de yaque (Leiarius marmoratus) durante el proceso de acostumbramiento a una dieta comercial, usando diferentes materias primas de origen animal como suplemento alimenticio.
Materiales y métodos
Localización
Los ensayos fueron realizados en el instituto de Acuicultura de la Universidad de los Llanos (IALL-UNILLANOS), localizado en el kilómetro 12 vía Puerto López, vereda Barcelona del Municipio de Villavicencio (Meta), ubicado a 418 m.s.n.m. Las condiciones climáticas de la estación piscícola son características de un clima tropical húmedo, con temperatura promedio de 25 °C, precipitación de 4050 mm anual y humedad relativa del 75%.
Material Biológico
Las larvas de las dos especies fueron obtenidas por reproducción inducida con Extracto de Hipófisis de Carpa (EHC) en las estaciones piscícolas Las Brisas y Langostinos del Llano, de los municipios de Acacias y Restrepo, respectivamente. Su alimentación exógena se inició a las 63 horas posteclosión (HPE), con presencia de un 10% de saco vitelino y con peso y longitud de 0.46 ± 0.2 mg y 4.7 ± 0.2 mm para L. marmoratus y 1.6 ± 0.32 mg y 6.8 ± 0.9 mm para Pseudoplatystoma sp., respectivamente.
Durante los primeros 15 días posteclosión (DPE), las larvas de L. marmoratus y Pseudoplatystoma sp. fueron alojadas en recipientes de acrílico, manteniendo una densidad de 10 larvas/L con el fin de realizar su acondicionamiento a las condiciones experimentales; durante este periodo fueron alimentadas con artemia salina, a razón de 750 nauplios/mL/larva por día.
Ensayos de acostumbramiento
Después del periodo de acondicionamiento a las condiciones experimentales, se tomaron al azar post-larvas de L. marmoratus y Pseudoplatystoma sp. de 16 DPE, con un peso y talla de 57.2 ± 4.8 mg y 17.1 ± 1.4 mm para L. marmoratus y de 55.9 ± 6.6 mg y 13.5 ± 2.7 mm para Pseudoplatystoma sp., respectivamente. De cada especie se seleccionaron al azar 240 larvas, las cuales fueron distribuidas en recipientes de acrílico con un volumen efectivo de 3L, manteniendo una densidad de 4 larvas/L. Cada recipiente fue considerado como una unidad experimental, las cuales fueron asignadas aleatoriamente a uno de 4 tratamientos, con cinco réplicas, así: tratamiento 1: alimento balanceado 40% de proteína bruta (C); tratamiento 2: alimento balanceado 40% de proteína bruta + corazón de bovino (CO); tratamiento 3: alimento balanceado 40% de proteína bruta + hígado (HI); tratamiento 4: alimento balanceado 40% de proteína bruta + hígado + aceite de pescado (HI+AP).
La proporción de los componentes de la dieta para cada tratamiento fue modificado semanalmente, iniciando con una proporción 80:20 de suplemento de proteína animal: alimento balanceado comercial y finalizando la sexta semana con 100:0, tal como se indica en la tabla 1. Las porciones de alimento balanceado y suplemento de proteína fueron mezcladas con gelatina sin sabor en proporciones iguales para cada tratamiento dentro del periodo de acostumbramiento, para otorgarle consistencia a la dieta suministrada y evitar su rápida desintegración y dilución en el agua. El suministro de la ración fue realizada ad - libitum 3 veces al día, a las 6:00, 16:00 y 21:00 horas. Durante las dos primeras semanas del ensayo, se suministró artemia salina en una o dos raciones (Tabla 1), con el fin de realizar la transición de alimento vivo a alimento húmedo.

Tanto en la fase de larvicultura como durante el periodo de condicionamiento progresivo fueron registrados diariamente la temperatura (°C), conductividad (μS/cm), oxígeno disuelto (mg/L), sólidos disueltos (TDS) y salinidad (PPT), empleando una sonda multiparamétrica YSI (MPS YSI 556, Spanish. qxp). Dos horas después de cada alimentación, se realizó recambio del 20% del agua del recipiente y retiro mediante sifonado de los residuos sólidos para evitar acumulación de materia orgánica y deterioro de la calidad del agua.
Se registró diariamente el número de larvas muertas, observando al estereoscopio posibles lesiones que explicaran la causa de su muerte, tales como mutilaciones que indicaran posibles agresiones consecuencia del canibalismo. Semanalmente se determinó el número de larvas desaparecidas, calculando la diferencia entre el número de larvas vivas al final de la semana menos las larvas muertas registradas durante la misma; este número se asumió como larvas consumidas totalmente a consecuencia del canibalismo intra-cohorte.
Evaluación del desempeño productivo
Para determinar el desempeño productivo a lo largo del ensayo, se realizaron 6 evaluaciones tomando el 20% de la población y registrando la longitud total (mm) y el peso corporal (mg) por medio de un calibrador (precisión de 0.01 mm) y balanza analítica (Ohaus, USA, 0.01 mg de sensibilidad), respectivamente.
Al finalizar la sexta semana se calculó la ganancia de peso [GP=(Peso final (mg) - peso inicial (mg)], la ganancia de talla [GT=(Talla final (mm) - Talla inicial (mm)], la tasa de crecimiento específico [TCE=(ln Peso final - ln Peso Inicial)/ 100], el factor de condición [Kn=(peso final (g) / longitud total (cm)] y la tasa de sobrevivencia [TS=(nº final de larvas x 100/nº inicial de larvas - nº de larvas)].
Análisis estadístico
Los datos fueron descritos como media ± error estándar de la media (ESM). Posteriormente, para analizar los efectos de los tratamientos se realizó un análisis de varianza (ANOVA) tipo MLG de dos factores (especies y fuentes de proteína), verificando previamente los supuestos de normalidad (prueba de Kolmogorov-Simirnov) y homogeneidad de varianzas (prueba de Levene). Los datos de sobrevivencia fueron transformados con la raíz del arco-seno. Se utilizó la prueba de Tukey para comparar las medias entre los diferentes tratamientos, como prueba a posteriori luego de establecer diferencias significativas. Se relacionó el peso y la longitud a través del tiempo mediante un análisis de regresión de la forma potencial para obtener la ecuación (peso= a longitud^b) con el fin de obtener la constante de crecimiento (b). El criterio de significancia fue p<0.05. Los análisis estadísticos fueron realizados usando el programa SPSS 13.0.
Resultados
Los parámetros de calidad de agua no presentaron variaciones entre los diferentes tratamientos a lo largo del ensayo (Tabla 2).

En general, se observaron diferencias significativas en el comportamiento de las variables productivas evaluadas entre post-larvas de L. marmoratus y Pseudoplatystoma sp., excepto para la tasa de crecimiento especifico (TCE), favoreciendo así el crecimiento de L. marmoratus. La ganancia de peso fue mayor en larvas de L. marmoratus comparado con Pseudoplatystoma sp. (F1, 39=372.8; p= 0.01), igualmente se observaron diferencias significativas por efectos del factor tipo de suplemento de proteína (F1, 39=372.8; p=0.01). El mejor desempeño productivo en términos de ganancia de peso corporal se observó en post-larvas que recibieron dietas con hígado + AP como suplemento de proteína para L. marmoratus (3213 ± 278 mg), mientras que para el Pseudoplatystoma sp. el mejor desempeño se observó con dietas que contenían corazón de bovino (695 ± 149 mg) (Figura 1a). En la interacción entre estos dos factores (especie x complemento de proteína) se observaron igualmente diferencias significativas 1,39=6.51; p=0.001), lo que permite establecer que ambas especies presentan respuestas diferentes de acuerdo con el tipo de suplemento de proteína utilizada para el acondicionamiento progresivo. En la ganancia de talla se observaron también diferencias significativas entre las especies (F1,39=349.9; p<0.001) siendo mayor en L. marmoratus (Figura 1b); para el factor suplemento de proteína utilizada en el acondicionamiento se encontraron diferencias significativas (F1,39=3.15; p=0.03), dando los mejores resultados en L. marmoratus el hígado + AP y corazón de bovino, mientras en Pseudoplatystoma sp. el mejor suplemento de proteína fue corazón de bovino; sin embargo, la interacción entre los dos factores no fue significativa (F1,39= 2.11; p=0.12).

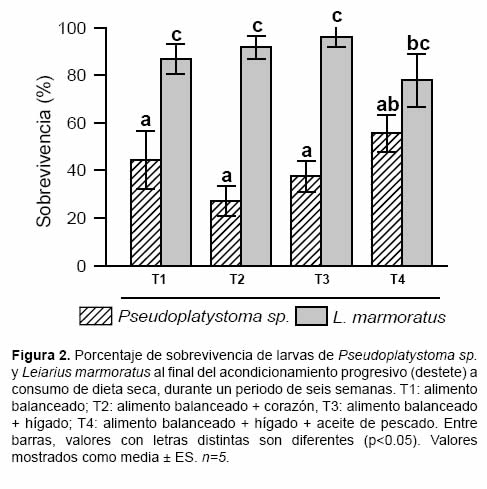
La sobrevivencia al fi nal del acondicionamiento presentó diferencias entre las especies (F1,39=45.23; p<0.001), siendo mayor para L. marmoratus (Figura 2). Aún cuando no se observaron diferencias significativas entre los suplementos proteicos utilizados (F1,39=0.33; p=0.801), la mayor sobrevivencia en L. marmoratus se observó en los tratamientos que incluyeron hígado (96 ± 2.2%), mientras que en Pseudoplatystoma sp. fue con hígado + AP (56 ± 7.7%); la interacción entre estos dos factores no presentó diferencias signifi cativas (F1,39=1.33; p=0.28).
La figura 3 muestra la mortalidad acumulada a lo largo del acostumbramiento a dieta seca para las dos especies, ilustrando como el canibalismo intra-cohorte (Figura 3 a,b) afecta en mayor proporción la sobrevivencia al final del ensayo en Pseudoplatystoma sp. En contraste, para L. marmoratus esta conducta se presentó en las dos primeras semanas en los tratamientos de alimento balanceado e hígado. Igualmente, la mortalidad asociada a factores diferentes al canibalismo afectó la sobrevivencia final para ambas especies especialmente para Pseudoplatystoma sp en el tratamiento alimento balanceado, por otro lado en L. marmoratus la mayor mortalidad no asociada al canibalismo se observó en el tratamiento que incluyó hígado + AP.

Igualmente, para el factor de condición (Kn) se observaron diferencias signifi cativas entre las especies (F1,39=121.28; p<0.001) (Figura 4), observándose los mayores valores para L. marmoratus; sin embargo, no se observaron diferencias entre los suplementos de proteína (F1,39=0.34; p<0.79), ni en la interacción entre estos dos factores (F1,39= 0.91; p=0.44).

(F1,39=1.36; p=0.25) (Figura 5), pero sí para los diferentes suplementos de proteína (F1,39=3.24; p=0.036). La interacción entre los dos factores no fue signifi cativa (F1, 39=2.79; p=0.057).

La figura 6 muestra la tendencia de la tasa de crecimiento especifi ca (TCE) a lo largo del ensayo, donde se observa para ambas especies una tendencia a disminuir el valor con el tiempo; sin embargo,
estos valores fueron mayores para L. marmoratus.

La relación del peso y la longitud para las dos especies presentaron ajustes significativos según la forma potencial de la forma peso = a*longitud b (Figura 7), sin embargo, los valores de la constante
b para Pseudoplatystoma sp fueron menores a 3, lo que indicaría un efecto negativo en la condición corporal a medida que aumenta su longitud, a diferencia de lo que se observó para larvas de L. marmoratus, en las cuales los valores son mayores a 3, observando para el tratamiento que recibió hígado + AP un valor de 4.5, refl ejando una muy buena condición.

Discusión
El acondicionamiento o destete consiste en el cambio gradual del suministro de alimento vivo a dieta artifi cial, proceso considerado como crítico en la crianza de peces carnívoros (Bock y Padovani, 2000; Kolkovski, 2001). El éxito de este proceso depende del alimento suministrado (digestibilidad y atractabilidad) y de las características de las post-larvas (edad, desarrollo del tracto digestivo)
(Dabrowski, 1984; Kolkovski, 2001); por lo tanto, un factor importante para el acostumbramiento exitoso es el tiempo mínimo de suministro de alimento vivo antes de iniciar el entrenamiento a consumo de dietas secas (Guerrero-Alvarado, 2003). En el presente estudio, el acondicionamiento de post-larvas de bagre rayado (Pseudoplatystoma sp.) y yaque (Leiarius marmoratus) se realizó entre los 16 y los 58 DPE, cuando se observaron los mejores resultados en cuanto a sobrevivencia, ganancia de talla y peso para post-larvas de L. marmoratus (Figura 1a y Figura 1b).
Los hábitos estrictamente carnívoros de Pseudoplatystoma sp. (Petrere et al., 2004) comparados con los hábitos omnívoros de L. marmoratus (Ramírez-Gil y Ajiaco-Martínez, 1997), favorecen en la primera el canibalismo durante el acondicionamiento a dieta seca (Figura 3a,b), evidenciado por la baja sobrevivencia de Pseudoplatystoma sp. al fi nalizar la sexta semana del ensayo (Figura 2), tal y como se registró a partir del conteo de larvas muertas con señales de agresión y registro de larvas desaparecidas, para las cuatro dietas ofrecidas (Figura 3a). Algunos trabajos reportaron en Pseudoplatystoma sp. (Nuñez et al., 2008; Díaz-Olarte et al., 2009) y L. marmoratus (Cruz-Casallas et al., 2008) que durante los primeros 15 DPE el canibalismo fue evidente en larvas alimentadas con artemia salina; este comportamiento disminuyó considerablemente durante el periodo de acostumbramiento para L. marmoratus, y se mantuvo para Pseudoplatystoma sp. (Figura 3b). Por lo tanto, el impacto del canibalismo depende directamente de la disponibilidad y de la calidad de los recursos alimenticios ofrecidos, ya que cualquier restricción alimenticia puede desencadenar o aumentar este comportamiento (Qin y Fast, 1996; Baras y Jobling, 2002), al igual que una descompensación o heterogeneidad en el crecimiento. Las dietas ofrecidas durante el ensayo, compuestas por una fracción de proteína de origen animal (corazón o hígado) y otra de alimento seco (alimento balanceado comercial), fueron más eficientes para cubrir las necesidades energéticas y de proteína para L. marmoratus, disminuyendo así su comportamiento caníbal.
En cuanto a las variables productivas de ganancia de talla y de peso corporal, factor de condición y tasa de crecimiento específico, el presente estudio concluye que el mayor rendimiento fue para post-larvas de L. marmoratus, comparado con Pseudoplatystoma sp. (Tabla 3). Estas diferencias en las respuestas al acondicionamiento progresivo entre las dos especies pueden explicarse por los estímulos químicos (olor, sabor y palatabilidad del alimento) y visuales (color, movimiento) que finalmente son la clave que resulte la comida (Kolkovski et al., 1997; 2000).
Algunos trabajos han discutido sobre los beneficios potenciales del alimento vivo como atrayente alimenticio para larvas de peces y se ha establecido que su uso durante los primeros estadios de vida mejora también el proceso de digestión, debido al aporte de enzimas digestivas (autolisis) y a la activación endógena de las mismas (Kolkovski et al., 1997; Kolkovski, 2001) y se sugiere que la actividad enzimática en el tracto digestivo, utilizada para digerir organismos vivos en los primeros estadios, es insuficiente para digerir una microdieta seca.
En el presente estudio, el suministro de Artemia salina durante las dos primeras semanas (Tabla 1), facilitó la digestión de la dieta de transición suministrada, lo cual se manifestó en la mayor ganancia de talla y peso corporal, factor de condición y sobrevivencia, como lo reportó Borges et al. (2006) para Piaractus mesopotamicus, en la cual encontraron que al utilizar conjuntamente organismos vivos y dieta inerte se obtienen mejores resultados en cuanto a parámetros productivos que cuando sólo se utilizaron organismos vivos, ya que el alimento vivo por sí solo no es suficiente para suplir los requerimientos nutricionales de las post-larvas de peces (Dabrowski, 1984).
El desarrollo del sistema digestivo en charácidos como Piaractus mesopotamicus permite realizar acondicionamiento progresivo a dietas secas a partir de los 10 DPE (Bock y Padovani, 2000), siendo menor que el tiempo necesario para silúridos como P. corruscans, para el cual Guerrero-Alvarado (2003) reportó un acondicionamiento exitoso y mejor aceptación al consumo de dietas secas en post larvas de 18 DPE sobre post-larvas de 13 DPE, evidenciando la importancia de la variable edad y tamaño corporal para el inicio del acostumbramiento en especies de hábitos carnívoros estrictos. Según los resultados obtenidos en el presente estudio, iniciar el proceso de acondicionamiento a consumo de dieta seca a los 15 DPE para larvas de L. marmoratus es adecuado, mientras que para Pseudoplatystoma sp. se recomienda iniciar el acondicionamiento a una edad superior a los 15 DPE.
El efecto de los diferentes suplementos proteicos utilizados durante el acondicionamiento progresivo presentó diferencias significativas entre las especies, así para Pseudoplatystoma sp. el mejor desempeño se observó en los tratamientos que incluyeron corazón de bovino, en cambio para
L. marmoratus fue hígado de bovino. El uso de estos suplementos proteicos de origen animal está vinculado con la activación directa e indirecta de enzimas como precursores o activadores de los procesos de asimilación y absorción, y por lo tanto, de la digestibilidad de las dietas secas (Kolkovski, 2001); siendo el principal inconveniente de las dietas formuladas que llegan a tener entre el 6090% de materia seca, mientras que las fuentes proteicas utilizadas están alrededor del 27% (Comunicación personal, Gutiérrez-Espinosa, Instituto de Acuicultura Universidad de los Llanos, 2009). Sustancias como neuropéptidos y factores de crecimiento que mejoran la digestión no son frecuentemente incluidas en las dietas formuladas; las fuentes proteicas de origen animal complementan eficientemente las dietas formuladas gracias a su aporte de aminoácidos principalmente. Los resultados obtenidos en el presente estudio mostraron como mejor suplemento de la dieta comercial el hígado de bovino, obteniendo resultados favorables en las variables productivas evaluadas (Figuras 3, 4 y 5).
A partir del proceso de acondicionamiento a consumo de dieta seca evaluado en el presente estudio se concluye que durante el mismo es necesario el uso de alimento vivo, así como de suplementos de proteína de origen animal que mejoren el factor de condición para las especies estudiadas. De igual forma, las especies pueden tener una respuesta diferente frente al acondicionamiento, debido posiblemente a sus diferencias en el desarrollo principalmente del tracto digestivo, por lo cual se hace necesario modificar los esquemas de acondicionamiento tradicionales, ajustando la edad de inicio del proceso y del cambio progresivo de dieta, acordes con estudios futuros relacionados con el desarrollo del tracto digestivo y de la actividad enzimática en cada especie.
Agradecimiento
Este artículo hace parte de los resultados del Convenio 015-03/06, entre el Ministerio de Agricultura y Desarrollo Rural y la Universidad de los Llanos.
Referencias
1. Aristizábal EO y Suárez J. Efficiency of co-feeding red porgy (Pagrus pagrus L.) larvae with live and compound diet. Rev Biol Mar Ocean 2006; 41:203-208.
[ Links ]2. Baras E, Jobling M. Dynamics of intracohort cannibalism in cultures fish. Aquacult Res 2002; 33:461-479.
[ Links ]3. Bock CL, Padovani CR. Considerações sobre a reprodução artificial e alevinagem de pacu (Piaractus mesopotamicus, Holmberg, 1887) em viveiros. Acta Sci 2000; 22:495-501.
[ Links ]4. Borges M y Portella M. Ingestão de ração e comportamento de larvas de pacu em resposta a estímulos químicos e visuais. Rev Bras Zootec 2006; 35:1887-1892.
[ Links ]5. Borges MT, Flores-Quintana CI, Carneiro DJ, Pizauro Junior JM, Portella MC. Suplementação de enzimas exógenas em dieta microparticulada para larvicultura do pacu. R Bras Zootec 2006; 35:2211-2218.
[ Links ]6. Cruz-Casallas NE, Diaz-Olarte JJ, Marciales-Caro LJ, Pabón-Peña FJ, Medina-Robles VM, Cruz-Casallas PE. Acondicionamiento a dieta seca de larvas de yaque (Leiarius marmoratus) obtenidas por reproducción artificial. Rev Colomb Cienc Pecu 2008; 21:482.
[ Links ]7. Dabrowski K. The feeding of fish larvae: present "state of the art" and perspectives. Reprod Nutr Develop 1984; 24:807-833.
[ Links ]8. Díaz-Olarte JJ, Cruz-Casallas NE, Marciales-Caro LJ, Medina-Robles VM, Cruz-Casallas PE. Efectos de la densidad de siembra y disponibilidad de alimento sobre el desarrollo y sobrevivencia de larvas de Pseudoplatystoma fasciatum. Orinoquia 2009;13:21-30.
[ Links ]9. Guerrero-Alvarado C. Treinamento alimentar de pintado Pseudoplatystoma coruscans (Agassiz, 1829): Sobrevivência, crescimento e aspectos económicos. Tesis de Maestría, Centro de Aqüicultura Jaboticabal; Universidade Estadual Paulista, São Paulo, Brasil 2003.
[ Links ]10. Hung L, Tuan N, Cacot P, Lazard J. Larval rearing of the Asian Catfish, Pangasius bocourti (Siluroidei, Pangasiidae): alternative feeds and weaning time. Aquaculture 2002; 212: 115-127.
[ Links ]11. Kerdchuen N, Legendre M. Larval rearing of an African catfish, Heterobranchus longifilis (Teleostei, Clariidae): a comparison between natural and artificial diet. Aquat Living Resour 1994; 7:247-253.
[ Links ]12. Kolkovski S, Arieli A, Tandler A. Visual and chemical cues stimulate microdiet ingestion in sea bream larvae. Aquac Int 1997; 5:27-536.
[ Links ]13. Kolkovski S, Czesny S, Dabrowski K. Use of krill hydrolysate as a feed attractant for fish larvae and juvenile. J World Aquac Soc 2000; 31:81-88.
[ Links ]14. Kolkovski S. Digestive enzymes in fish larvae and juveniles - implications and applications to formulated diets. Aquaculture 2001; 200:181-201.
[ Links ]15. Nuñez J, Duque R, Corcuy-Arana N, Duponchelle F, Renno J, Raynaud T. Induced breeding and larval rearing of Surubí, Pseudoplatystoma fasciatum (Linneaus, 1766), from the bolivian Amazon. Aquac Res 2008; 39:764-776.
[ Links ]16. Petrere M, Borges R, Agudelo E, Corrales B. Review of the large catfish fisheries in the upper Amazon and stock depletion of piraiba (Brachyplatystoma filamentosum Lichtenstein). Rev Fish Biol Fish 2004; 14:403- 414.
[ Links ]17. Qin J, Fast A. Size and feed dependent cannibalism with juvenile snakehead Channa striatus. Aquaculture 1996; 144:313-320.
[ Links ]18. Ramírez-Gil H y Ajiaco-Martínez R. Aspectos preliminares de la biología pesquera del Yaque, Leiarius marmoratus (Gill, 1870) (Pisces: Siluriformes: Pimelodidae) en la parte alta del río Meta (Orinoquia Colombiana). Boletin científico INPA 1997; 5: 75-87.
[ Links ]19. Segura L, Hayashi C, de Souza S, Soares C. Canibalismo entre larvas de pintado, Pseudoplatystoma corruscans, cultivadas sob diferentes densidades de estocagem. Acta Sci 2004; 26:299-302.
[ Links ]20. Sevilla A, Günther J. Growth and feeding level in pre-weaning Tambaqui Colossoma macropomum Larvae. J World Aquac Soc 2000; 31:218-224.
[ Links ]21. Vergara-Morelo R, Hoyos-Pacheco J. Evaluación del entrenamiento del bagre blanco (Sorubim cuspicaudus littmann, burr & nass, 2000) al consumo de dietas secas. Tesis de pregrado, Programa de acuicultura; Universidad de Córdoba, Montería, Colombia 2005.
[ Links ]
