











 English (pdf)
English (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mail Cited by SciELO
Cited by SciELO  Cited by Google
Cited by Google  Similars in
SciELO
Similars in
SciELO  Similars in Google
Similars in Google 
 Permalink
PermalinkIntroduction
Sheep breeding is exploited on all continents, with wide dissemination mainly due to its ability to adapt to different climates, terrain and vegetation. Mutton production experienced 27% growth in Brazil over the last 20 years, despite the reduced demand for wool (MAPA, 2013).
The increase in mutton consumption has required greater production efficiency. Therefore, it is necessary to improve the management of herds through detailed characterization of breeds, breeding systems and critical phases, taking into account the different production regions. Nutritional and health problems are the main cause of the limited efficiency of ruminant production (Hernandez et al., 2013). During the peripartum period, depression of the immune system is directly related to the physiological state of the animal (Silva et al., 2003 to Nieto et al., 2003) due to hormonal changes that occur near lambing and during lactation (Gennari et al., 2002;.Aya et al., 2008). These changes are called "the peripartum phenomenon" (Amarante et al., 2004; Rocha et al., 2004).
Nutrients and energy requirements increase during pre-natal and early lactation, concomitant with a reduction in dry matter intake due to rumen compression by the fetus and production of hormones involved in pregnancy (Rodrigues et al., 2006). This nutritional challenge is also associated with increased susceptibility to gastrointestinal parasites. An increase in the number of nematode eggs per gram of feces in sheep during peripartum can occur due to inadequate supply of dietary protein (Donaldson et al., 1998 and 2001; Houdijk et al., 2001).
Although a number of studies have investigated the association between nutrition and gastrointestinal infections in sheep (Hoste et al., 2005) few have focused on the potential variations of hematological, biochemical and parasitological parameters associated with peripartum, a critical stage for productive and reproductive efficiency. This knowledge is necessary to improve management practices, particularly because of the large heterogeneity of values within this species due to differences of breed, gender, age, nutrition, management systems (Kerr, 2003) and physiological status (Cardoso et al., 2010).
In this study, we characterized the hematological, biochemical and parasitological values in the peripartum period in Santa Ines sheep, the main native breed used in Brazil and one of great interest in other South American countries.
Materials and Methods
Ethical Considerations
This study was conducted and approved by the ethics committee for animal research at Instituto de Zootecnia (finding no. 142/2011).
Local
This study was conducted at Centro de Pesquisa e Desenvolvimento de Zootecnia Diversificada, at Instituto de Zootecnia, of Agência Paulista de Tecnologia dos Agronegócios (APTA), da Secretaria de Agricultura e Abastecimento, located in Nova Odessa, São Paulo state (22°42’ S and 47°18’ W).
According to the Köppen classification, the climate in the Nova Odessa region is CWa, characterized as tropical with relatively dry and hot summers and rainy winters. Average rainfall is 1,317 mm per year and the temperature is between 10 ° C and 35 ° C, with an average of 26 ° C. The average annual relative humidity is approximately 75%.
Animals and management
A total of 70 Santa Ines ewes kept under a natural breeding system were tested for pregnancy by ultrasound. Of these, we used 45 sheep diagnosed with close gestational age and 50 kg average body weight. Sheep were kept under rotational grazing on Panicum maximum Jacq. cv. Aruana and cv. Tanzania pastures, with ad libitum access to water and mineral mixture. During late pregnancy, sheep were confined in paddocks and supplemented with corn silage ad libitum and 0.300 kg concentrate (16% crude protein). They remained in the paddocks until the lambs were weaned at 3 months of age. Feed samples were taken every 15 days and frozen at -20 ºC for later analysis.
Sample collection and evaluation of animals
Blood and stool samples were collected every 14 days during five months, for a total of 10 collections. At this time, sheep were also weighed and evaluated for body condition score and conjunctival staining. Blood samples were also collected for hematological and biochemical analyses on the day of lambing. Body condition score was determined by palpation of the lower back (Sanudo and Sierra, 1986) using a half-point scoring (Pugh, 2004). Parasitological and hematological analyses were performed at the Laboratory for Fecal Analysis and Laboratory for Clinical Analysis, respectively, of Instituto de Zootecnia de Nova Odessa/APTA.
Parasitological examinations
Stool samples were collected directly from the rectum for egg counting (eggs per gram of feces; EPG) according to the modified technique by Gordon and Whitlock (1939). Fecal culture of infective larvae was conducted for identification of parasite genra (Roberts and O’Sullivan, 1950).
Hematological analysis
Blood samples from all animals were collected by venipuncture of the jugular vein in Vacutainer tubes (5 ml) containing ethylenediaminetetraacetic acid (EDTA). Packed cell volume analysis (PVC) was conducted after microhematocrit centrifugation. Hemoglobin (Hb) concentration was measured by the cyanmethemoglobin method using a semi-automatic biochemical analyzer (Bio 200 L Bioplus) using commercial kits for hemoglobin determination (Labtest). Total plasma protein (TPP) was determined by the refraction technique. Total leukocyte count (Tot Leuk) and erythrocyte count (RBC) were determined manually in a Neubauer hemocytometer (Schalm and Carroll, 1986). Tests identified blood smears and differential count of segmented neutrophils (Seg Neut), lymphocytes (Lymph), eosinophils (Eosin), monocytes (Mono), basophils (Baso) and rods (Bast) (Schalm and Carroll, 1986). No rods were observed, so this parameter was removed from the statistical analysis.
Biochemical analysis
The blood samples were subjected to the following biochemical analyses: globulin, albumin, serum protein (Ser Pt), urea, creatinine, total bilirubin (BT), direct bilirubin (BD), indirect bilirubin (BI), glucose (Gluco), cholesterol, AST (AST), calcium, iron (Fe), and phosphorus. The tests were performed in a Bio 200L semiautomatic biochemical analyzer (Bioplus, São Paulo, Brazil) using commercial kits (Labtest, Lagoa Santa, Minas Gerais, Brazil).
Statistical analysis
The assumption of normal distribution of all variables was verified. The variables fibrino gen, total leukocytes, segmented neutrophils, lymphocytes, eosinophils, monocytes, tricho strongylid, Strongyloides spp, Moniezia spp and Eimeria spp which did not meet the assumption of normality were transformed. The data were transformed for the FAMACHA and fibrinogen variables by raising to the -0.2 power. Square roots were used for total leukocytes and erythro cytes, and the other non-normal variables were log10 transformed. The means are presented in the original scale.
Analysis of variance (ANOVA) was conducted using the PROC GLM routine of SAS for all variables (processed or not) considering repeated measures. The model and analysis considered the effect of weight as a covariate in addition to the effects of treatment and time and their interaction. The means were compared by the Tukey test considering 5% probability for Type I error.
Results
Feeding and performance
Sheep ate a daily average of 1.85 kg dry matter and 177 g crude protein at the beginning of the experiment, with consumption decreasing in the peripartum month (September) and increasing linearly until the end of lactation (November), when it reached 2 kg dry matter and 200 g crude protein (Figure 1).

Body weight increased significantly from the beginning of the study until lambing, when it was the highest. Subsequently, there was a sharp and significant drop in weight from day 1-13 after birth, when body weight returned to a value similar to that observed >30 days before birth. A further reduction in weight was also observed from days 14-27 (lactation), after which it remained constant. The analysis of BCS showed a significant reduction associated with the peripartum period, which was particularly accentuated during the peak of lactation (up to 83 days after birth). From day 84 onwards, BCS was not different from that during pregnancy (Table 1).
Parasitological analysis
Table 2 shows the results of the endoparasite analysis. The mean FAMACHA score varied from 1.2 (1-13 days) to 1.7 (<-30 days), with no difference between periods (p>0.05). Trichostrongylid FEC was, however, significantly higher during gestation and peripartum than during lactation, with a particularly sharp reduction in FEC towards the end of the latter period.
Table 1 Least square means of body weight and body condition score (BCS) of Santa Ines ewes.
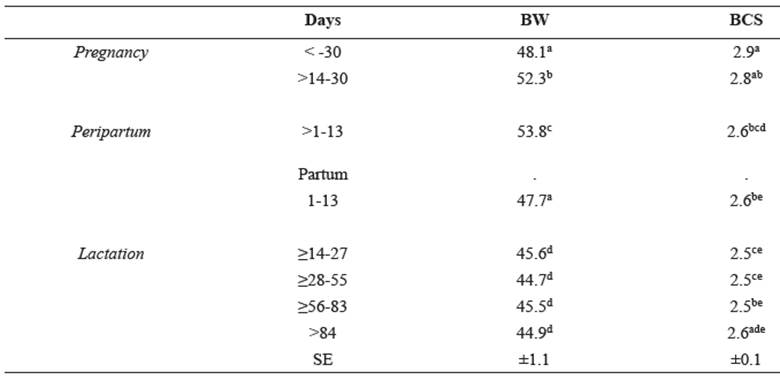
Different superscript letters (a, b, c, d, e) within the same column indicate significant differences at p<0.05.
Table 2 Least square means of FAMACHA© (± standard error) and parasitological variables of Santa Ines ewes (± standard error).
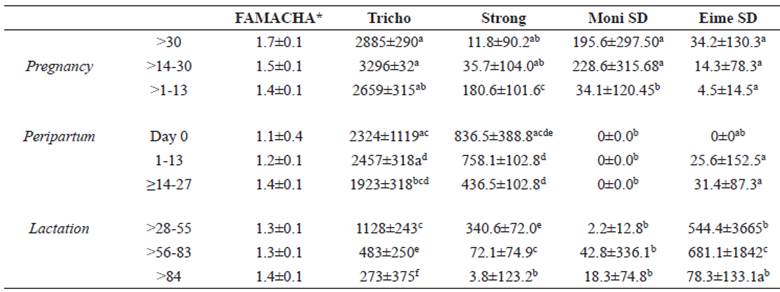
*In those cases where ER, SE were used. Otherwise, different superscript letters (a, b, c, d, e) within the same column indicate significant differences at p<0.05 (Tukey-Kramer). Thrichostrongylides (Tricho), Strongyloides spp (Strong), Moniezia spp (Moni), Eimeria spp (Eime).
Unexpectedly, Trichostrongylid FEC was not significantly higher on the lambing date. Strongyloides spp. FEC increased from gestation to birth, with accentuated increase on the birth date, and subsequent reduction afterwards. The Moniezia spp. count showed the highest values in the first two gestational periods, while Eimeria counts had the lowest values in the first six periods (pregnancy, peripartum and lactation up to 27 days postpartum), differing from later lactation periods.
The coproculture results showed average counts of Trichostrongylidae, which is the parasite genus of greatest interest, with predominance of Haemonchus ssp (92.85%).
Hematological profile
The hematological variables showed significant variations across the gestational period (Table 3). As expected, there was a gradual increase in packed cell volume over the course of the study, and a decrease in mean corpuscular volume (MVC). The lowest values of hemoglobin concentration (Hb), total plasma protein (TPP), and mean corpuscular hemoglobin concentration (MCHC) were observed near the birth date.
This date was also associated with the highest values of leucocytes, neutrophils and lymphocytes (Table 4). Eosinophils and monocytes were, conversely, particularly low at birth.
Blood biochemistry
All blood biochemistry analyses, regarding metabolites, enzymes (Tables 5 and 6) and minerals (Table 7), were performed in a Bio 200L semiautomatic biochemical analyzer (Bioplus, São Paulo, Brazil), with Labtest commercial kits (Labtest, Lagoa Santa, Minas Gerais, Brazil). Blood levels of metabolites, globulin, albumin, total serum protein, urea, creatinine, glucose (Table 5) total bilirubin, direct bilirubin, indirect bilirubin, cholesterol and AST differed significantly (p <0.05) with the different physiological conditions (Table 6). The globulin values ranged from 3.88 (≥14-27 days) to 4.85 (> 84 days), with a significant increase over the course of lactation. The values of albumin and serum protein were significantly lower in the birth date, although similar to the average associated with day 84 onwards. The values of urea (Table 5) decreased from gestation to peripartum, subsequently increasing over the lactation period. There were no clear trends in the variation of creatinine values.
Sheep had lower levels of total, direct and indirect bilirubin on lambing day (p>0.05) compared to all other periods, except for the direct bilirubin, which was similar to that of the period > 84 days postpartum (Table 6).
Cholesterol and AST had constant variations during all periods, with respective amplitudes of 48.2 ± 1.8 (1-13 days) to 60.8 ± 1.8 (> 1-13) and 82.6 ± 3.7 (≥14-27) to 111.0 ± 4.4 (> 84).
The serum levels of calcium, phosphorus and iron showed significant differences (p<0.0001) between the periods of pregnancy, birth and lactation. The mean changes in calcium, phosphorus and iron were independent of the physiological condition.
Table 3 Mean (± standard error) of hematological variables of Santa Ines ewes, according to physiological stage.
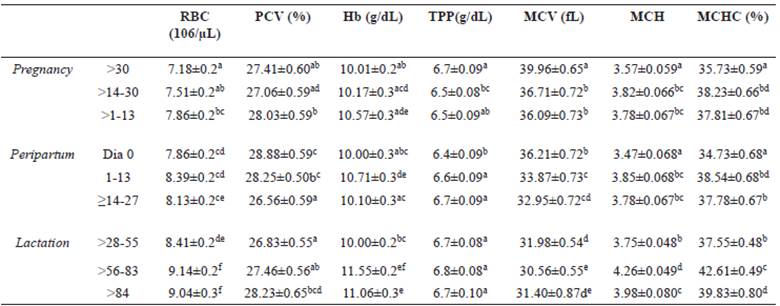
Average followed by different superscript letters (a, b, c, d, e) within the same column differ by the Tukey’s test (p<0.05). (RBC) erythrocytes, (PCV) packed cell volume, (Hb) hemoglobin concentration, (TPP) total plasm protein, (MCV) mean corpuscular volume, (MCH) mean corpuscular hemoglobin, (MCHC) mean corpuscular hemoglobin concentration.
Table 4 Least square mean (± standard error) of fibrinogen (Fibri), total leukocytes (Tot Leuk), segmented neutrophils (Seg Neut), lymphocytes (Lymph), eosinophils (Eosin), and monocytes (Mono) of Santa Ines ewes according to physiological stage.
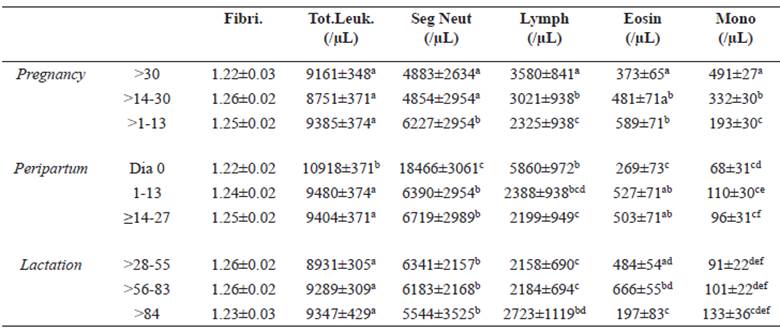
Different superscript letters (a, b, c, d, e) within the same column indicate significant differences at p<0.05 (Tukey-Kramer).
Table 5 Mean (± standard error) of globulin, albumin, serum protein (Ser Pt), urea, creatinine (Creatinin), and glucose (Gluco) of Santa Ines ewes according to physiological stage.
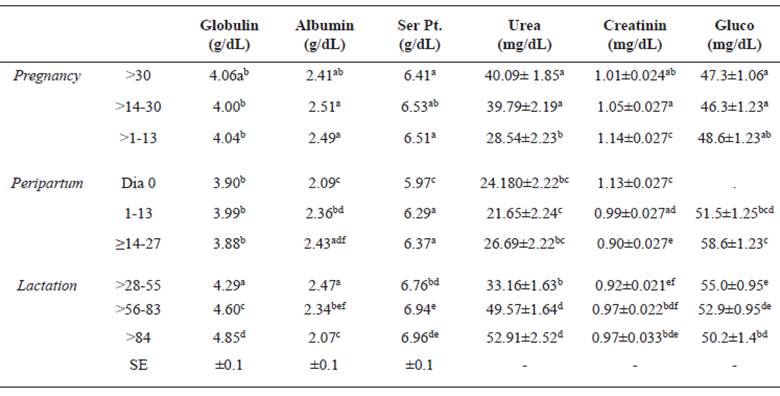
Different superscript letters (a, b, c, d, e) within the same column indicate significant differences at p<0.05 (Tukey-Kramer).
Table 6 Mean (± standard error) of total bilirubin (BT), direct bilirubin (BD), indirect bilirubin (BI), cholesterol, and AST of Santa Ines ewes according to physiological stage.
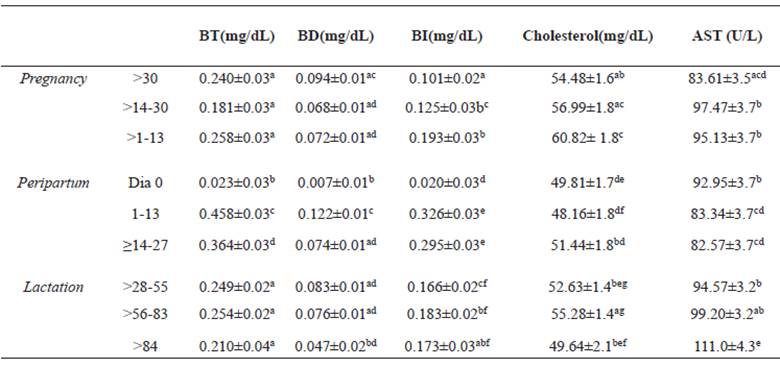
Different superscript letters (a, b, c, d, e) within the same column indicate significant differences at p<0.05 (Tukey-Kramer).

Discussion
Previous research suggests that feed intake of sheePrevious research suggests that feed intake of sheep and other ruminants is often insufficient in the final period of gestation and early lactation. Our results show that although a decrease in dry matter and crude protein intake occurred in these stages, consumption was sufficient to meet the animals’ requirements in these physiological states (NRC, 2006) in confined conditions. Accordingly, we observed relatively small weight and body condition score variations.
Although the peripartum period is often associated with increased susceptibility to gastrointestinal parasitic infections, there were no significant variations in trichostrongylid counts between gestational and peripartum periods. Haemonchus spp. genus are highly pathogenic blood-suckers, causing a decrease in the packed cell volume and hence increasing the ocular mucosa coloration score (FAMACHA). Haemonchus spp. had high mean prevalence (92.85%), while Trichostrongylid egg counts decreased during the experiment. The FAMACHA score was constant during different physiological conditions. It is unlikely that this low association between Trichostrongylid FEC and the FAMACHA score resulted from a subjective assignment of this score since the association between PCV and FAMACHA was high. Therefore, the weak association between FAMACHA score and Trichostrongylid count may have resulted from the fact that sheep were already infected before the experiment. Although the Trichostrongylid FEC was high until the peripartum period and decreased with the onset of lactation, the appropriate protein intake might have allowed maintenance of PCV values predominantly within the range considered normal for sheep (27 to 45%; Pugh, 2005) throughout the experiment. The only exception was a small decrease in PCV just after the beginning of lactation, but was followed by swift recovery to values within the normal range. Other parasites with lower pathogenicity than Haemonchus spp. (Afonso et al., 2013) were also identified, such as Trichostrongyloidea (Trichostrongylus, Cooperia and Oesophagostomum), Strongyloides spp., Eimeria spp. and Moniezia spp. Moniezia and Eimeria FEC were higher in the pregnancy and lactation periods, respectively, probably due to the management used. Better food, confined animals are less exposed to infection by Moniezia (albeit more exposed to infection by Eimeria due to trough feeding). Although Haemonchus spp. can cause severe anemia and hypoproteinemia (Faria Junior et al., 2002), depression, loss of body condition, reduced productivity, and even death (Kaplan et al., 2004), no animal died or had to be dewormed before or during our experiment.
As expected, there was a variation in the hematological profile of animals between periods. The values of hematological variables were predominantly below the normal range for the species, possibly due to the negative action that hormone variation has on hematological variables (Kerr, 2003). Hematological parameters change easily and fast during peripartum (Cardoso et al., 2010) and the PCV decreases as pregnancy progresses due to the physiological hemodilution in this period (Brito et al., 2006). Recent studies suggest that even healthy Santa Ines sheep have lower erythrocyte, hemoglobin and PCV levels and higher MCHC values (Salviano et al., 2013) compared to the reference values for the species found in the literature (Schalm, 1981; Harvey and Meyer, 1998; Pugh, 2005). Our results showed increased amounts of red blood cells and hemoglobin from pregnancy until the end of lactation. However, we did not observe lower PCV values during gestation, as reported in the literature (Brito et al., 2006). We also found higher MCHC values in relation to the literature, including those suggested by Salviano et al. (2013) for non-pregnant sheep. These findings reinforce the hypothesis that breed can significantly affect hematological patterns, and emphasizes the need for studies on specific breeds considering age, sex and physiological condition to avoid misinterpretation of results.
During pregnancy, hormone action on the immune system (Kerr, 2003) is necessary to suppress the defense mechanisms and thus promote survival and development of the fetus (Lloyd, 1983). The low total leukocyte levels observed before delivery may be related to this factor, while the high level on the day of delivery may be related to the migration of these cells into the uterus, where there is tissue injury caused by birth.
Segmented neutrophils act as a first line of defense against infection. Many researchers have investigated the activity of these leukocytes close to delivery (Guidry et al., 1976; Fonteque et al., 2013). The level of segmented neutrophils increased as delivery approached, peaking on day 0 (delivery), with a gradual decrease afterward. This subsequent reduction may be related to the migration of these cells to injury sites, corroborating the results of Fontequete et al. (2013), who observed decrease in total leukocytes and segmented neutrophils during postpartum. However, they also noted increased segmented neutrophils and total leukocytes after delivery, indicating the likely need to replenish the number of circulatory cells due to their flow to the uterus. We did not observe this pattern, probably because sheep were parasitized, which kept the immune system active.
The observed decrease in lymphocytes and monocytes before delivery, starting 30 days beforehand and declining sharply 15 days before delivery, may have occurred because of migration of cells to the uterus and or abomasum. Migration of leukocytes to the abomasum occurs to enable antigen recognition of the nematode, followed by stimulation of the immune system to generate the appropriate response to fight parasites (Douch, 1990). The activation of larvae, previously in hypobiosis, happens due to immunosuppression in response to circulating cortisol in the days before delivery. The number of EPG also increased in these same periods. Lymphocyte concentration was lower during postpartum than on the delivery day, whereas monocytes showed higher values 15 days after delivery and after more than 84 days of lactation. The high score in the first period after delivery may be related to peak lactation and attempt of the body to restore immunity after the stress period caused by constant hormonal changes.
Eosinophil counts increased gradually until the day of delivery, decreased considerably in the delivery day, and then rose gradually in the postpartum period until 84 days in milk, after which the concentrations declined. These results are similar to those reported by Fonteque et al. (2013) who found that stress caused by birthing in goats is overcome in the first week after delivery, with more stable counts, a fact confirmed in our study. However, these findings contrast with those of Sabag et al. (1978) who noted a decreasing number of circulating eosinophils due to the action of cortisol associated with inhibition of eosinophilopoiesis in the marrow and/or migration of these cells to other tissues.
The globulin profile behaved proportionally to the nutritional requirement in each physiological condition, with a progressive increase from the peak of lactation (≥14- 27 days) until the end of lactation, when the demand decreased due to lower milk production. Moreover, our results showed a decrease of serum albumin and protein values in the peripartum period, specifically on the day of birth, with subsequent increase during lactation. This decline can be expected to occur starting in the last third of gestation until peak lactation, due to three main factors. First, there is a decrease in consumption of dry matter and crude protein in late pregnancy and early lactation. Second, there is a larger requirement of the fetus in this stage, with 70% of growth happening in the last 50 days of pregnancy (Russell, 1979). Finally, there is higher nutritional requirement until peak milk production (Houdijk, 2008), with changes in hormone concentrations to promote gluconeogenesis and mobilization of adipose tissue to provide energy to the fetus and the mammary glands (Rodrigues et al. 2007). However, the decrease in serum albumin and protein levels occurred only on the day of delivery, although food consumption (DM and CP) apparently met the animals’ requirements, as demonstrated by maintenance of body condition throughout the experiment. The specific reasons for the reduction in these parameters only on the lambing day remain to be determined.
The urea profile is directly related to protein intake and renal function; changes in its level indicate possible kidney disorders.
As expected, the urea values found in this study followed the protein intake pattern in the different physiological stages, with decrease from gestation to peripartum periods and subsequent increase in the lactation period. Variations in creatinine were not related to these periods and were always below the normal range for the species (1.2 to 1.7 mg/dL).
Calcium values increased from the gestation peripartum. There was a decrease in the level of this mineral during peak milk production (≥14-27) in relation to peripartum, which might be related to milk production requirements. However, endocrine control of calcium means calcium is not a good nutritional indicator, although this mineral is closely associated with metabolism in the peripartum period (Gonzalez, 2000). The small variation in serum phosphorus profile was not related to the physiological condition. However, in some stages of gestation, peripartum and lactation, values lower than the normal ones for other sheep breeds were noted (5.1 to 7.3 mg/dL, Gonzalez et al., 2006). In the postpartum period, this mineral -which is an indirect indicator of energy metabolism- can be used in the form of adenosine triphosphate, which would cause a decrease in its concentration (Feijó et al., 2014).
According to Borjesson et al. (2000), glucose concentration in ruminants is readily altered with stress and excitement due to catecholamine release. Since the assessed periods can be considered stressful, especially peripartum, the changes noted can be considered normal. However, ruminants have an excellent mechanism for maintenance of glucose, keeping it at relatively constant concentrations. This makes observations of large changes unlikely, because only very severe dietary restrictions would lead to hypoglycemia (Peixoto et al., 2010).
An increase in AST levels is associated with liver and/or muscle damage. However, our results were within the normal range for the species and in accordance with those reported by Avellanet et al. (2007) for the Xisqueta breed (66.0 to 194.0 U/L). Similarly, increased concentration of blood bilirubin is most commonly caused by hemolysis or liver diseases (Raposo et al., 2004). Batista et al. (2009), studying healthy sheep in two age groups (3-5 years and over 5 years), found mean values of total bilirubin of 0.046 and 0.037 mg/ dL, direct bilirubin of 0.020 and 0.016 mg/dL, and indirect bilirubin of 0.026 and 0.021 mg/ dL, respectively. However, according to Pugh (2005), normal values for the total and direct bilirubin are between 0.1 to 0.5 mg/dL and 0.0 to 0.27 mg/dL, respectively. Thus, despite the variation found in different physiological periods, just total bilirubin on the delivery day had a lower average than what is considered normal for the species.
There was little variation in cholesterol levels throughout the experiment, with lower values in the peripartum and at the end of lactation -when the animals were already at a lower nutritional level to promote drying of milk. Ribeiro et al. (2004) found no difference in cholesterol levels between empty and pregnant ewes and suggested that blood metabolites do not vary with the reproductive status of well-fed sheep. However, Peixoto et al. (2010) reported that formation of steroid hormones during gestation causes part of the cholesterol in the blood to be reabsorbed, decreasing its level at that stage.
Mendonça et al. (2005) reported that serum changes of some minerals, including iron, can result from infectious diseases of ruminants, including mastitis. Kaneko et al. (1997) considered serum iron values between 166 and 221 mg/dL to be normal for the species. Despite the variations in periods, the concentrations remained within the normal range.
The results of this study indicate that the peripartum period exerts an influence on the hematological, parasitological and biochemical parameters of sheep. Nonetheless, correct knowledge and interpretation of physiological parameters during this phase can prevent mistaken diagnosis and provide adequate support during this stage.

