











 Spanish (pdf)
Spanish (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mail Cited by SciELO
Cited by SciELO  Cited by Google
Cited by Google  Similars in
SciELO
Similars in
SciELO  Similars in Google
Similars in Google 
 Permalink
Permalink
INTRODUCCIÓN
La Orinoquía es reconocida como un área estratégica debido a su variedad de ecosistemas naturales y transformados. Teniendo en cuenta la clasificación de Bustamante (2019) , se divide en siete paisajes fisiográficos con características diferentes en cuanto a clima, origen geológico, hidrología, suelos, vegetación natural y uso de la tierra. El piedemonte de esta área está ubicado en un rango altitudinal de 300-1100 m y corresponde a una subunidad de paisaje de elevada importancia biogeográfica, ya que se encuentra en una zona de transición entre la montaña de la Orinoquía y la sabana (Trujillo & Henao, 2018).
El piedemonte llanero tiene una gran relevancia natural en comparación con el resto de la Orinoquía; es considerado como uno de los paisajes con mayor productividad primaria (Briceño-Vanegas & Gallego-Herrera, 2023). Sus bosques poseen una multiplicidad de condiciones abióticas, como su geomorfología, riqueza hídrica y complejidad biogeográfica, que propician el establecimiento de una elevada diversidad de especies (Díaz-Pérez et al., 2018). Por otra parte, la presencia de estos bosques permite proteger las fuentes hídricas de las subcuencas, que son cruciales en la región, además de otros servicios ecosistémicos como el almacenamiento de carbono, el hábitat para la fauna y el sostenimiento del suelo (Pardo-Rozo, 2022).
La biodiversidad presente en los ecosistemas de bosque de piedemonte está siendo amenazada por diferentes procesos que involucran, entre otros, las actividades humanas y el cambio climático (Etter et al., 2017). Aunque las características topográficas de estos bosques no son las ideales para la agricultura y la ganadería, han sido explotados de manera continua para estos fines (Díaz-Pérez et al., 2018). Esto ha ocasionado la pérdida de una gran parte de la cobertura de los bosques de piedemonte, lo que pone en peligro la prestación de servicios ambientales por parte de las comunidades vegetales allí establecidas (Rodríguez-Ovalle & Paéz-Vásquez, 2022).
Debido a las diferentes amenazas descritas, el objetivo de este estudio fue caracterizar, a nivel florístico y estructural, un bosque de piedemonte ubicado en la vereda Aguas Claras del municipio de Cubarral, Meta, Colombia. Esta información puede ser la línea base para futuros estudios sobre el manejo y la conservación de estas áreas.
MATERIALES Y MÉTODOS
Área de estudio
Este estudio se llevó a cabo en el noroccidente del departamento del Meta, en la vereda Aguas Claras del municipio de Cubarral, entre los 702 y 795 m de altitud. Cubarral posee un área total de 1532 km², su temperatura anual varía entre 18 y 30 °C, la precipitación media anual es de 5121 mm (Rodríguez-Ovalle & Paéz-Vásquez, 2022) y, según la clasificación de zonas de vida de Holdridge (1987) , corresponde a un bosque tropical muy húmedo (bmh-T). En la actualidad, la fuente de ingresos más representativa del municipio es la ganadería, aunque los ingresos de algunos habitantes provienen de la tala indiscriminada en ecosistemas vulnerables.
Muestreo de la vegetación
La fase de campo se realizó entre julio y diciembre del año 2019. Se establecieron diez transectos de 50 x 2 m, distribuidos al azar, evitando que se sobrepusieran, y distanciados entre sí por lo menos en 20 m, para un total de 0.1 ha (Mendoza, 2008). En cada transecto se censaron y se recolectaron todos los individuos con un diámetro a la altura del pecho (DAP) mayor o igual a 2.5 cm, teniendo en cuenta los estándares de herborización propuestos por Forero (1997). Se registró la circunferencia a la altura del pecho (CAP) a 1.3 m del suelo (Mendoza, 2008) y la altura total de cada individuo recolectado. Además, se georreferenció cada uno de los transectos, tanto al inicio como al final, y se realizó un registro fotográfico.
Determinación taxonómica
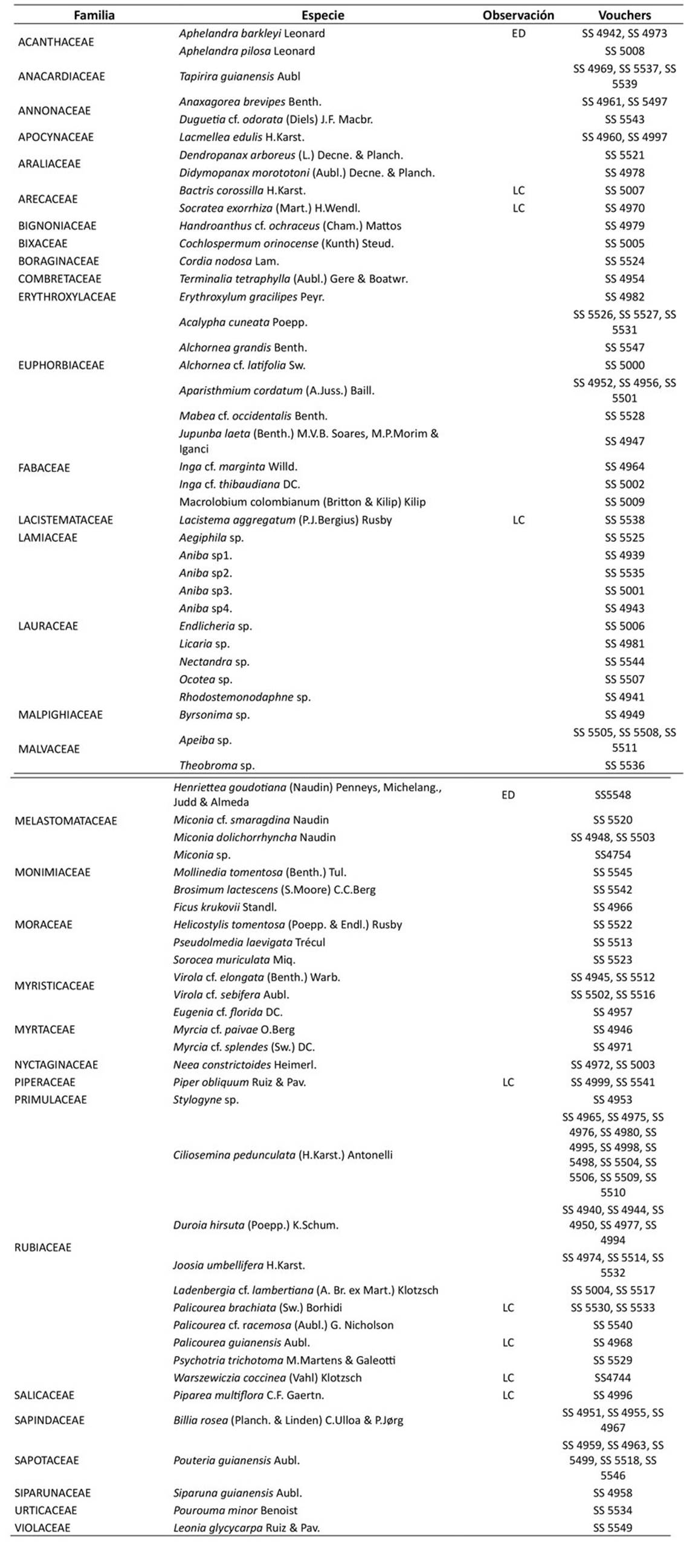
Para la determinación taxonómica, se revisaron las colecciones del Herbario LLANOS y las colecciones botánicas de los herbarios virtuales COAH, UDBC y COL, además de literatura especializada (Gentry, 1993; Quiñones, 2001; Mendoza et al., 2004). La nomenclatura botánica se basó en el sistema de clasificación APG IV (Chase et al., 2016), y se validaron los nombres científicos aceptados en la base de datos Plants of the World Online (POWO, 2023). Las colecciones botánicas fueron depositadas en el Herbario LLANOS después del proceso de determinación.
Análisis de la información
Composición florística
Para procesar la información, se tuvieron en cuenta la composición florística y los parámetros estructurales. En cuanto a la composición florística, se elaboró un listado de las especies presentes en el área de estudio, organizado según la literatura específica para cada una de las familias.
Estructura vertical y horizontal
Con respecto a la caracterización estructural, se realizó una distribución de individuos por clases de altura y DAP, por medio del software R versión 4.2.1 (R core Team, 2023) y el paquete fdth (Faria et al., 2023) considerando la fórmula propuesta por Sturges (1926) . Con base en estos cálculos, se estableció la estructura horizontal y vertical de la vegetación, obteniendo las clases diamétricas y altimétricas en las que se distribuyen los diferentes individuos del área muestreada.
Valores de importancia ecológica
Para caracterizar la importancia ecológica de cada especie dentro de la comunidad vegetal se tuvieron en cuenta los valores relativos de la densidad (número de individuos por especie/número total de individuos) x 100, la dominancia (sumatoria de las áreas basales de todos los individuos de una especie/sumatoria de las áreas basales de todas las especies) x 100 y la frecuencia (número de transectos en las que se encuentra una especie/número total de transectos) x 100 (Rangel & Velásquez, 1997; Mostacedo & Fredericksen, 2000).
Con base en la sumatoria de los anteriores parámetros, se determinó el índice de valor de importancia ecológica (IVI) (Rangel & Velásquez, 1997). Por último, se calculó el índice de valor de importancia para familias (IVF) con la sumatoria de los valores relativos de la densidad, la dominancia y la riqueza de cada familia (Robinson et al., 2003).
RESULTADOS
Composición florística
Se registró un total de 288 individuos, agrupados en 71 especies, 59 géneros y 33 familias. Las familias con mayor riqueza se observan en la Tabla 1. Dentro de estas, se destacaron Lauraceae (9 spp.) y Rubiaceae (9 spp.). Los géneros que presentaron mayor número de especies fueron Aniba (4 spp.), Miconia (3 spp.) y Palicourea (3 spp.). Por otro lado, 50 de los géneros registrados en este estudio estuvieron representados por una sola especie (Anexo 1).
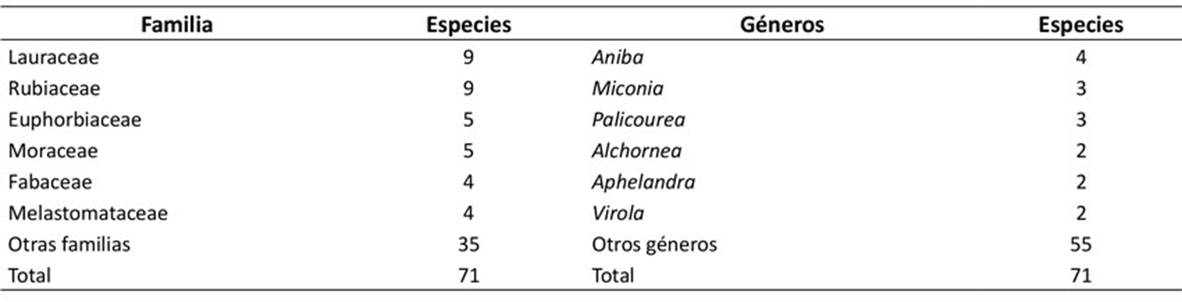
Valores de importancia ecológica
Las familias con mayor importancia ecológica (IVF) fueron Rubiaceae (142.8 %), Lauraceae (18.0 %), Euphorbiaceae (15.3 %), Arecaceae (15.1 %) y Moraceae (9.7 %). Las especies con mayores valores de importancia ecológica fueron Ciliosemina pedunculata (H. Karst.) Antonelli (62.6 %), Warszewiczia coccinea (Vahl) Klotzsch (39.4 %), Socratea exorrhiza (Mart.) H.Wendl. (23.5 %), Duroia hirsuta (Poepp.) K.Schum. (16.1 %) y Pouteria guianensis Aubl. (12.4 %) (Tabla 2).
Estructura vertical y horizontal
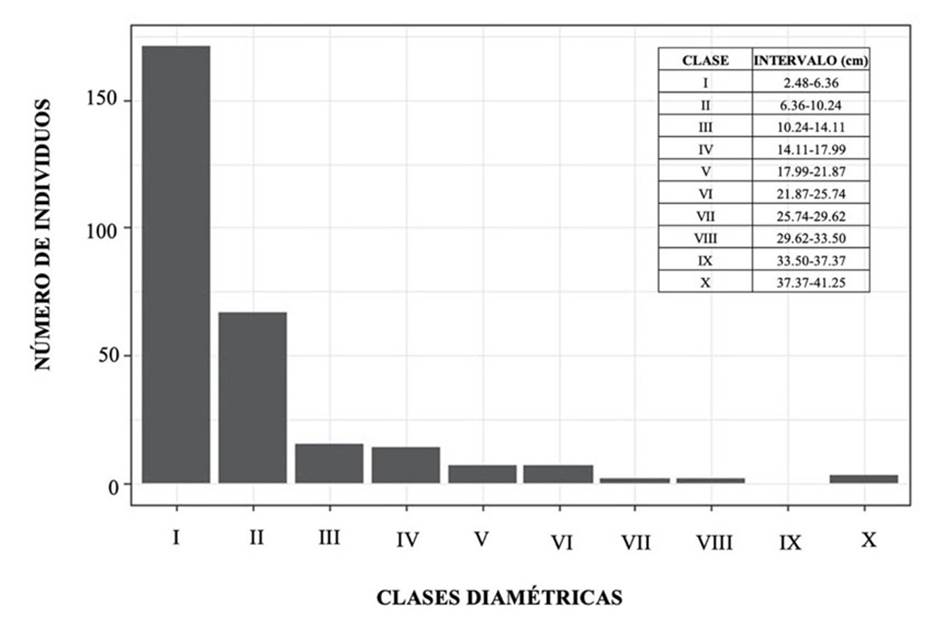
La vegetación presentó un predominio de clase de altura en los dos primeros intervalos (1.58 a 5.16 m y 5.16 a 8.74 m). Allí se encontró la mayor parte de individuos (180) (Figura 1). Respecto a las clases diamétricas, se observó un comportamiento en forma de ‘jota’ invertida: el mayor número de individuos (171) se concentró en la categoría más baja (2.58 a 6.36 cm), disminuyendo de manera progresiva conforme los valores del diámetro incrementaban (Figura 2).
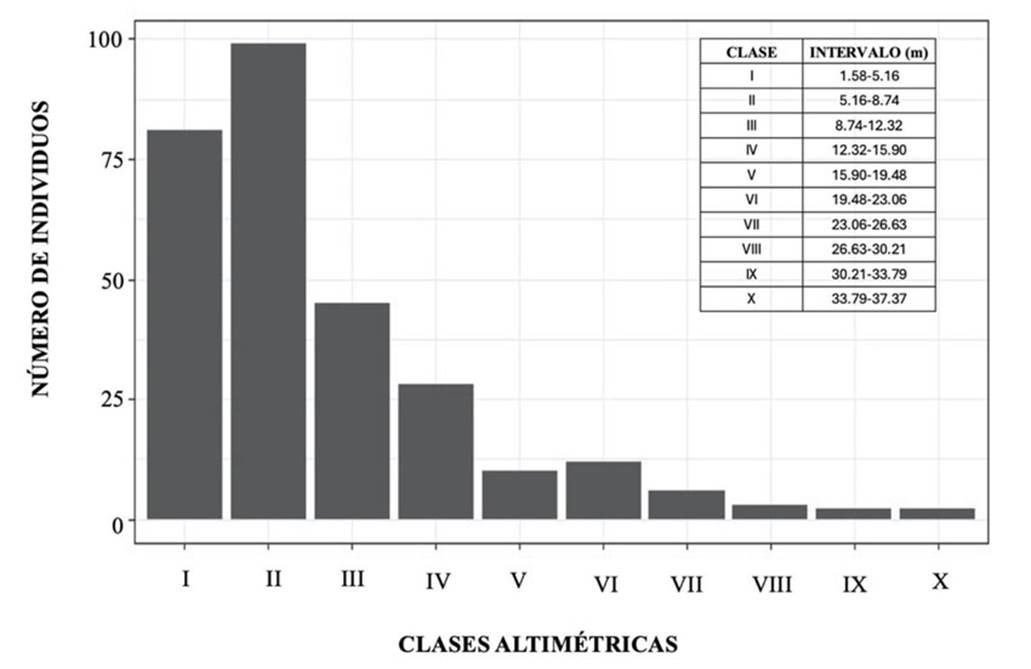
Figura 1 Distribución de árboles y arbustos por clases de altura en un bosque de piedemonte de Cubarral, Meta
DISCUSIÓN
Composición florística
La riqueza florística en cuanto a especies, géneros y familias fue menor a la de los resultados de otros estudios en los bosques de la Cordillera Oriental de Norte de Santander (Dueñas et al., 2007) y en los bosques del piedemonte llanero en Casanare (Díaz-Pérez et al., 2018) y Meta (Cárdenas-Torres, 2014; Carvajal et al., 2012). Estas diferencias pueden estar relacionadas con los muestreos, pues se establecieron ocho parcelas de 20 x 50 m en los trabajos de Cárdenas-Torres (2014) y Díaz-Pérez et al. (2018), y dos parcelas permanentes de 100 x 100 m en Carvajal et al. (2012).
El hallazgo de las familias Rubiaceae y Lauraceae como las más representativas en este estudio concuerda con lo reportado para los bosques de piedemonte en la Cordillera Oriental (Dueñas et al., 2007; Carvajal et al., 2007; Carvajal et al., 2012). Estos resultados son similares a los obtenidos por Cárdenas-Torres (2014) y Díaz-Pérez et al. (2018) , pero difieren en tanto registran a Fabaceae como la de mayor número de especies, taxón considerado como el más diverso en bosques de piedemonte (Trujillo & Henao, 2018). A nivel de géneros, solo se comparte a Miconia como el de mayor riqueza con los otros estudios previos (Dueñas et al., 2007; Cárdenas-Torres, 2014; Díaz-Pérez et al., 2018).
Valores de importancia ecológica
El IVF se ve fuertemente influenciado por la familia Rubiaceae, que presenta elevados valores en cuanto a dominancia y densidad. Esto se debe a que es un taxón típico de los bosques secundarios tropicales (López-Herrera, 2022) y posee tasas altas de germinación como estrategia de vida, lo que propicia procesos ecológicos que intervienen en la regeneración natural de los hábitats perturbados (Sánchez et al., 2019).
Algunas de las familias reportadas con altos valores de importancia ecológica se han registrado en otros bosques de piedemonte de la Cordillera Oriental. Aunque en un orden diferente, en los bosques del Parque Nacional Natural Catatumbo Barí se comparten las familias Rubiaceae, Lauraceae, Arecaceae y Moraceae (Dueñas et al., 2007). Asimismo, en los bosques de Aguazul (Díaz-Pérez et al., 2018), la familia Rubiaceae también se reporta entre las de mayor IVF, pero con un valor notablemente menor (142.8 vs. 30 %).
Los valores de IVI más altos fueron registrados por dos especies de la familia Rubiaceae (C. pedunculata y W. coccinea), lo que ratifica su importancia en estos ecosistemas. La especie C. pedunculata ha sido registrada como un taxón con un alto valor de importancia ecológica en estudios previos de la zona (Carvajal et al., 2007). Además, en los bosques fragmentados de Acacías y Guamal solo se comparten las especies S. exorrhiza y D. hirsuta (Cárdenas-Torres, 2014). Esto puede estar vinculado con el rápido crecimiento en claros y bordes de bosque que presenta la primera (Richards et al., 2022) y la asociación con hormigas de la segunda, las cuales posiblemente estén destruyendo plántulas de otras especies, como se ha documentado en otras áreas (Frederickson et al., 2005; Fernández et al., 2019).
Estructura vertical
Las clases altimétricas presentan un comportamiento similar a una distribución normal sesgada hacia las alturas menores, en donde el intervalo II (5.16 a 8.74 m) es el que agrupa la mayor cantidad de individuos (99). Las distribuciones de alturas en otros bosques de piedemonte presentan una forma de ‘jota’ invertida (Dueñas et al., 2007) y campana de Gauss (Carvajal et al., 2012). Estas diferencias con el bosque estudiado pueden estar asociadas a la etapa de sucesión, siendo esta más avanzada en los bosques que presentan una tendencia unimodal (Cabrera-Amaya & Rivera-Díaz, 2016). Con respecto a las alturas, Carvajal et al., (2012) no registran ningún individuo con una altura > 25 m, contrario al presente estudio, donde se reportan, en el estrato arbóreo superior (>25 m), un total de once individuos correspondientes a nueve especies. Por otra parte, Dueñas et al., (2007), registran que la mayor cantidad de individuos se concentra en alturas de entre 1.3 y 4.9 m, y solo nueve superan los 23.5 m.
Estructura horizontal
La mayor parte de los individuos (82.6 %) se distribuyen en las clases diamétricas I y II (2.48 a 6.36 cm y 6.36 a 10.24 cm respectivamente) y decrecen drásticamente en los intervalos superiores. Esta disposición es afín con lo reportado en bosques de la misma zona de estudio (Carvajal, 2012), donde las gráficas han mostrado una tendencia en forma de ‘jota’ invertida. El predominio de una gran cantidad de árboles en las categorías inferiores puede indicar que el bosque se encuentra en un proceso de regeneración temprana (Cuéllar-Cardozo et al., 2022); los individuos registrados pueden ser las primeras especies que se establecen después de una alteración y podrían estar aún en crecimiento o son especies de corto periodo de vida que no alcanzan grandes diámetros y que son abundantes en este componente de regeneración natural (Quinteros-Gómez et al., 2021).
CONCLUSIONES
La composición florística del bosque de Cubarral coincide con lo reportado en otros bosques de piedemonte, donde tanto las familias más representativas como las especies con mayor importancia ecológica desempeñan un papel importante en la dinámica ecosistémica. La estructura también coincide con la de otros bosques de piedemonte: la mayor parte de los individuos se concentra en los primeros intervalos de altura y diámetro, en respuesta a un proceso de recuperación natural en el área. Se recomienda realizar un muestreo más representativo que permita establecer la verdadera diversidad de este ecosistema, ya que, con el diseño de muestreo implementado, se obtuvieron bajos niveles de riqueza en comparación con otros trabajos en el piedemonte llanero.

