










Services on Demand
Journal
Article
 Spanish (pdf)
Spanish (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mailIndicators
-
 Cited by SciELO
Cited by SciELO -
 Access statistics
Access statistics
Related links
-
 Cited by Google
Cited by Google -
 Similars in
SciELO
Similars in
SciELO -
 Similars in Google
Similars in Google
Share

 Permalink
PermalinkActa Medica Colombiana
Print version ISSN 0120-2448
Acta Med Colomb vol.30 no.1 Bogotá Jan./Mar. 2005
Luis Miguel Gómez: Estudiante de Maestría, Unidad de Biología Celular e Inmunogenética, Corporación para Investigaciones Biológicas, y Facultad de Biología, Universidad de Antioquia, Medellín; Dr. Carlos Cañas: Jefe de Reumatología, Fundación Clínica Valle del Lili, Cali, Colombia; Dr. Juan-Manuel Anaya: Profesor Titular, Unidad de Biología Celular e Inmunogenética, Corporación para Investigaciones Biológicas, Universidad del Rosario, Medellín.
Correspondencia al Dr. Juan-Manuel Anaya, MD. Corporación para Investigaciones Biológicas (CIB), Cra. 72 A No. 78 B 141, Medellín, Colombia, Tel: (4) 441 0855, 441 8846, Fax: (4) 441 5514 - E-mail: janaya@cib.org.co
Recibido: 10/09/04. Aceptado: 28/03/05
Resumen
Los receptores Fc, miembros de la superfamilia de las inmunoglobulinas, participan en fenómenos inflamatorios comoanti-inflamatorios, influyendo sobre la inmunidad innata y adquirida. Factores ambientales y genéticos afectan su expresión. Los receptores Fc participan en el desarrollo de autoinmunidad y otras patologías como cáncer y enfermedades infecciosas. En autoinmunidad cumplen un papel protagónico, ya que controlan una serie de funciones inmunológicas que incluyen reacciones mediadas por complejos inmunes, liberación de citoquinas, lisis celular dependiente de complemento, apoptosis, degranulación de mastocitos, endocitosis y potenciación de la presentación antigénica clase I y clase II. La deficiencia de FcgRIIb está asociada con una susceptibilidad a desarrollar enfermedades autoinmunes. Igualmente, el polimorfismo de los genes para estos receptores se asocia a varias enfermedades autoinmunes. En el presente artículo se revisan las principales funciones de los receptores Fc, su polimorfismo genético y su implicación clínica, en particular en enfermedades autoinmunes.
Palabras clave: receptores Fc, autoinmunidad, genética.
Abstract
Fc receptors (FcR) belong to the immunoglobulin super-family and have been proposed as important regulators of inflammation, acting in both innate and acquired immunity. Environmental and genetic factors affect FcR expression. FcR receptors participate in the pathogenesis of autoimmunity, cancer and infectious diseases. In autoimmunity, they play important roles in different settings including immune-complex reactions, cytokines release, complement-dependent cellular citotoxicity, apoptosis, mastocyte-degranulation, endocytosis, and antigen presentation enhancement. FcgRIIb deficiency has been associated to the development of autoimmunity. Likewise polymorphism in the FcR genes is a susceptibility risk for autoimmune pathology. This article reviews the main functional aspects of FcR and their genetic polymorphism as well as their practical impact in the clinic, with emphasis in autoimmune diseases.
Key Words: Fc receptors, autoimmunity, genetics
Introducción
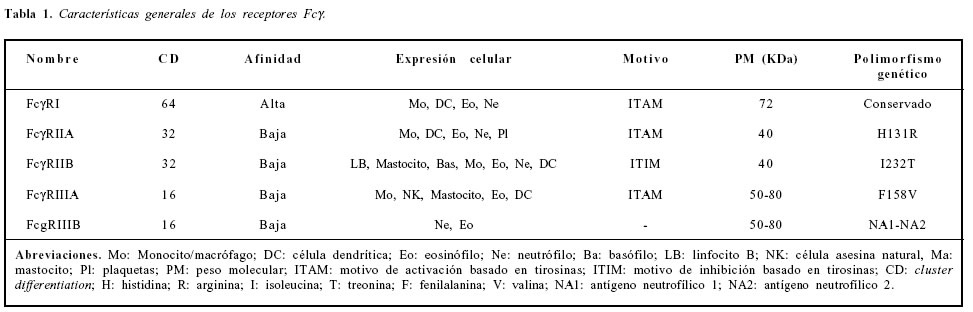
Los receptores Fc son una familia heterogénea de glicoproteínas de membrana que unen la fracción Fc de la inmunoglobulina G (IgG)(1). Han sido clasificados en tres tipos básicos: FcgRI, FcgRII y FcgRIII (Tabla 1). FcgRI posee alta afinidad por la IgG en forma monomérica, mientras que FcgRII y FcgRIII, son de baja afinidad y sólo unen la IgG en forma de complejos inmunes. Dentro de las funciones efectoras importantes de estos receptores se pueden encontrar la fagocitosis y la liberación de citoquinas pro-inflamatorias. Dichos receptores fueron descubiertos hace treinta y cinco años cuando se observó que las inmunoglobulinas IgG eran citofílicas para macrófagos en presencia de glóbulos rojos opsonizados, independiente de la región que se unía al péptido (región Fab) y dependiente de la región de la fracción cristalizable conocida como Fc (2).
Los receptores Fc poseen una función importante en el sistema inmune ya que proporcionan la conexión entre inmunidad humoral y celular. La unión de estos receptores con su ligando, como la IgG, produce cambios conformacionales y por lo tanto activación de funciones efectoras como citotoxicidad dependiente de anticuerpos, ingesta de complejos inmunes, entre otras. Por otro lado, un subtipo de receptores Fc, que contienen una secuencia particular de aminoácidos en el dominio intracitoplasmático, modula la inhibición de la activación celular y es un mecanismo común para receptores que transmiten una señal negativa.
Las fisiopatologías de las enfermedades autoinmunes son el resultado de la pérdida de tolerancia con una exagerada activación de linfocitos autorreactivos, producción de autoanticuerpos y una inflamación constante en los sitios de depósito de complejos inmunes. Dado que en las enfermedades autoinmunes se observa una inhibición de la respuesta tanto humoral como celular, es fundamental determinar si los receptores Fc desempeñan un papel importante en estas patologías. En el presente artículo se revisan las principales funciones de los receptores Fc, su polimorfismo genético y la relación con patología autoinmune. Al final se presenta un glosario con las principales abreviaciones utilizadas en el mismo y su definición biológica.
Estructura molecular
Los receptores Fc están compuestos por una cadena principal alfa (a) y unas cadenas accesorias como beta (b), gama (g), xi (x), que influyen sobre la respuesta inmunológica, debido a los receptores de activación o de inhibición que posean, ya sea en su cadena a o en sus cadenas accesorias, mencionadas anteriormente (3). Los receptores Fc contienen tres dominios clasificados de acuerdo con su ubicación, llamados dominio extracitoplásmático, dominio transmembrana y dominio de activación/inhibición. El receptor de activación posee una región basada en tirosinas (ITAM) en su región citoplasmática que actúa como unidad central para la transmisión de señales, que redunda en funciones como citotoxicidad dependiente de anticuerpos (ADCC), citotoxicidad dependiente de complemento (CDCC), endocitosis, regulación de anticuerpos, producción de citoquinas inflamatorias, potenciación de la presentación antigénica y regulación del estallido respiratorio (4,5). El receptor Fc inhibidor contiene un dominio de inhibición basado en tirosinas (ITIM) en su porción citoplasmática, que regula las funciones de los dominios activadores como es el caso de la regulación de anticuerpos y la activación de la apoptosis de las células B (6). Los receptores Fc son clasificados también por la afinidad del receptor por la IgG. Por lo tanto existen los receptores de alta afinidad y de baja afinidad (Tabla 1) (7).
Funciones
Éstas pueden clasificarse en tres: regulación de la respuesta inmune, ingesta de complejos inmunes y funciones del receptor neonatal para IgG (FcRn).
Dentro de la primera función se pueden destacar la proliferación de las células B, fagocitosis por macrófagos y degranulación de mastocitos. La internalización, después de la unión receptor-inmunoglobulina, da lugar a varias funciones efectoras como fagocitosis, histolisis, degranulación y activación de sitios blancos en regiones génicas de unión a factores de transcripción, las cuales inician cascadas de inflamación. Sin embargo, cuando se da la unión del receptor inhibidor FcgRIIb, se regulan de una manera negativa dichos receptores, expresándose ubicuamente en células del sistema inmune e inhibiendo funciones específicas como activación, proliferación de células B y degranulación de mastocitos. El sistema ITAM/ITIM se ha consolidado como un mecanismo del sistema inmunológico para regular sus actividades.
En la segunda función, la ingesta de complejos inmunes, los FcgRs pueden activar la internalización de antígenos capturados por anticuerpos, lo cual finaliza con la degradación del complejo antígeno-inmunoglobulina-complemento, definición de la ruta de procesamiento y presentación antigénica ya sea mediada por moléculas HLA clase I o clase II. Es importante recalcar que en enfermedades como el lupus eritematoso sistémico (LES) existen fallas en la eliminación de los complejos inmunes (8). Hallazgos recientes han puntualizado sobre la eficiencia de la presentación antigénica si la ingesta de los complejos inmunes es mediada por receptores Fc (9).
La tercera función está relacionada con el receptor Fc neonatal (FcgRn). Éste fue identificado primero en ratones como el receptor que transfería gamaglobulinas (IgGs) de la madre al hijo vía intestino neonatal (10). Sin embargo, se ha comprobado que el FcgRn es importante también para el mantenimiento de los niveles séricos de IgG (11). La expresión del FcRn en tejidos como hígado, glándula mamaria e intestino adulto podría hacer pensar en que estos receptores modulan el transporte de IgG a estos sitios. Esta hipótesis es reforzada por la afinidad del FcRn por la IgG y transportarse así por los tejidos (12).
Mecanismos de señalización celular
La activación o inhibición de receptores Fc depende de la presencia de ITAM o ITIM respectivamente, ya sea de una manera intrínseca en el receptor, como en el caso de FcgRIIa, o como parte de una cadena accesoria g o x como el caso de FcgRIIIa y FcgRI. La excepción es el FcgRIIIb el cual no posee porción citoplasmática y, por lo tanto, no contiene dominios ITAM (ver glosario) (13).
Activación
FcgRI y FcgRIII hacen parte de una familia de proteínas llamadas receptores inmunes reconocedores de cadenas múltiples (MIRR), los cuales se presentan en forma de complejos hetero-oligoméricos de una cadena alfa que une la inmunoglobulina y otra cadena de señalización. Los dominios citoplasmáticos de los MIRR son de Tipo ITAM, es decir, de activación (14). Por lo tanto cuando se une una IgG a FcgR se produce un cambio conformacional que conlleva a la activación de una familia de tirosin kinasas llamadas Src, como lo son Hck, Lyn y Fyr (15). Posteriormente, se produce el agrupamiento de moléculas con dominios SH2, como la kinasa Syk, que se unen a ITAM fosforilado, como una especie de anclaje molecular (Figura 1). Es importante señalar que dependiendo del tipo celular y la clase de receptor Fc usado, pueden estar involucradas diferentes clases de kinasas. Posteriormente, se procede la activación de la kinasa IP3, la cual desencadena la producción de PIP3 y a su vez la unión de moléculas con dominios PH como PLCg (16) y otra serie de kinasas como Tec dentro de las cuales se incluiyen btk, itk y emt, que son expresadas en diferentes tipos de células mieloides (17). La fosfolipasa C del subtipo gama (PLCg) da lugar a dos componentes: diacilglicerol (DAG) y el 1,4,5-trifosfato (IP3). DAG permanece unido a membrana e IP3 promueve el paso continuo de calcio (18).
Inhibición
Las respuestas de inhibición en la señalización celular están dadas por los receptores que poseen dominios tipo ITIM, como el FcgRIIb.
El receptor inhibidor FcgRIIb se une a IgG con baja afinidad e interactúa con complejos inmunes sólo a pH fisiológicos (19). La unión de complejos inmunes entre el receptor de la célula B (BCR) y FcgRIIb promueve la actividad inhibitoria que se lleva a cabo con el bloqueo en el flujo de calcio y por lo tanto reducción de la proliferación y diferenciación de células B, afectando la secreción de inmunoglobulinas (20).
ITIM se diferencia de ITAM en un residuo hidrofóbico pequeño en la posición 2 que generalmente se encuentra justo antes de la tirosina en el motivo ITIM dando lugar a inhibición más que activación (21).
La cascada de señalización se inicia con la unión del receptor Fc con la inmunoglobulina, generando un cambio conformacional que activa una kinasa de la familia SRC, llamada Lyn. Esta modificación desencadena el agrupamiento de fosfatasas que contiene dominios SH2 como SHP1, SHP2 y fosfatasas que contienen inositol llamadas SHIP (22). Esta modificación que se realiza en el dominio SH2, que es el sitio de unión para la fosfatasa SHIP, impide la activación de ITAM por hidrólisis de PIP3, que es una molécula que participa en la cascada de activación. En ausencia de PIP3, las proteínas de unión a dominios PH como Btk y PLCg, son liberados de la membrana y la señal de entrada de calcio a la célula es bloqueada (23) (Figura 2).
Resumiendo, las consecuencias de la transmisión de señales después de la unión receptor Fc con la inmunoglobulina son, en el caso de activación, la entrada de calcio a la célula y en el caso de la inhibición, el bloqueo de ésta. El calcio tiene una función importante en este tipo de respuesta, ya que funciones como la ADCC, CDCC, fagocitosis, liberación de citoquinas e inflamación, son dependientes de calcio, es decir, si no hay la suficiente concentración de calcio en la célula, dichos procesos no son posibles.
Genética
Los genes que codifican para los receptores de baja afinidad (FcgRIIa, b, c y FcgRIIIa y b) hacen parte del complejo de receptores Fc y están ubicados en la región 1q23. Su estudio ha sido difícil por la alta homología, fruto de duplicaciones y recombinaciones en esta región. En un sentido centrómero-telómero se ubican así: FcgRIIA, FcgRIIIA, FcgRIIC, FcgRIIIB, FcgRIIB, en una región de 10 Kb y con direcciones de inicio diferentes de la transcripción (Figura 3) (24). Esta es una región que ha sufrido rearreglos en sus genes, como es el caso de FcgRIIA y FcgRIIB, que dieron lugar, por entrecruzamiento desigual (entrecruzamiento entre cromosomas homólogos que no están perfectamente emparejados) a FcgRIIC (25). Además, es frecuente el "rearreglo alternativo" (formación de nuevos exones a partir de los preexistentes, cuando se da el proceso de transcripción de ADN), que produce diferentes formas solubles de la proteína (Figura 3).
El FcgRIIA, presente en mononucleares, neutrófilos y plaquetas, tiene dos alelos funcionalmente diferentes y expresados en forma codominante, que se generan del cambio de una guanina por una adenina, conocidos como H131 y R131, los cuales se diferencian en un aminoácido en la posición 131 del segundo dominio extracelular, el cual es sitio de unión para el fragmento Fc de la IgG (26). Este cambio genera diferentes afinidades por el tipo de inmunoglobulina, siendo especialmente la isoforma que contiene histidina la que se une más específicamente con IgG2, dando lugar a cambios funcionales importantes, ya que la IgG2 es un mal activador de la vía clásica del complemento (26, 27).
FcgRIIb presenta un cambio de isoleucina por treonina en la región transmembrana de la proteína siendo de importancia funcional, ya que es el único receptor de esta familia que tiene funciones de inhibición de la respuesta inflamatoria (28).
FcgRIIIa posee una sustitución de timina por una guanina, resultando en un cambio en el aminoácido valina por fenilalanina. Los pacientes que tienen el genotipo valina/valina son considerados buenos respondedores para la unión de IgG1, IgG3 e IgG4 (29).
FcgRIIIb posee una sustitución de cuatro aminoácidos denominada el antígeno neutrofílico (NA), que corresponde al dominio distal en la porción extracitoplasmática del receptor. Esta porción es un sitio importante que afecta la glicosilación de la proteína (30). Así, las variantes alélicas son denominadas NA1 en el caso de ser el genotipo sin la sustitución y NA2 para la sustitución. La fagocitosis de complejos inmunes inducida por el alelo NA1 es más eficiente que por el alelo NA2 (Tabla 1) (31).
Fc y autoinmunidad
Las enfermedades autoinmunes, desde el punto de vista etiológico, conjugan factores medioambientales, genéticos, inmunológicos y hormonales. Éstas se caracterizan por la presencia de una respuesta linfocitaria T y/o B autorreactiva en ausencia de alguna causa discernible (infección o cáncer), acompañados de inflamación, producción de autoanticuerpos, presencia de autoantígenos (modificados o no), pérdida de la tolerancia y daño tisular. Desde el punto de vista inmunogenético, las enfermedades autoinmunes son complejas, debido a que no siguen un patrón de herencia mendeliano y son poligénicas, sumado a interacciones génicas como epístasis y desequilibrio de ligamiento (32). El estudio de los receptores Fc en el contexto de autoinmunidad ha despertado un gran interés por su implicación biológica en la depuración de complejos inmunes, entre otras. Dentro del componente genético, estudios en ratones que son deficientes para FcgRs, han mostrado una incapacidad para fagocitar complejos inmunes de una manera eficiente (33). Además estos ratones sufren espontáneamente varias enfermedades autoinmunes (34).
Los polimorfismos de los receptores Fc influyen en la eficacia de la respuesta celular y han sido asociados con enfermedades inflamatorias, infecciosas y autoinmunes, así como con la severidad de éstas. Además, estudios recientes han demostrado que estos polimorfismos también pueden afectar la respuesta a diversos medicamentos biológicos como los anticuerpos monoclonales (35). Desequilibrios entre el balance de receptores Fc inhibidores o activadores puede ocasionar una ruptura de la tolerancia y desencadenar enfermedad autoinmune (Figura 4).

Un resumen de los principales estudios de polimorfismos genéticos de los receptores Fcg en enfermedades autoinmunes es dado en la Tabla 2. Algunos polimorfismos en las regiones reguladoras se han asociado con enfermedad. Finalmente, el uso de agentes terapéuticos que modulan la expresión de dichos receptores pueden ser útiles en la práctica clínica (Tabla 3). Una regulación exagerada en el receptor inhibidor podría evitar reacciones de defensa tan importantes como la ADCC, fagocitosis y liberación de citoquinas (Figura 5).

Glosario
ADCC: citotoxicidad celular dependiente de anticuerpos.
Btk: kinasa perteneciente a la familia Tec, denominada tirosin kinasa de Bruton.
CDCC: citotoxicidad celular dependiente de complemento.
DAG: molécula de señalización que se queda unida a membrana, generada por la fosfolipasa Cg. Su principal función es activar una enzima llamada protein kinasa C.
Emt: kinasa perteneciente a la familia de las Tec, que son expresadas en diferentes tipos de células mieliodes.
Fyr: kinasas de la familia SRC
HcK: kinasas de la familia SRC
ITAM: inmunorreceptor con motivos de activación basados en tirosinas. Son motivos compuestos de dos copias, con la secuencia tirosina-X-X-leucina (donde la X es cualquier aminoácido) que se encuentran en la cola citoplasmática, de varias proteínas del sistema inmune.
ITIM: inmunorreceptor con motivos de inhibición basados en tirosinas. Es un motivo de seis aminoácidos, isoleucina-X-tirosina-X-X-leucina (donde X es cualquier aminoácido), que se encuentra en la cola citoplasmática de varios receptores inhibidores del sistema inmune.
Itk: kinasa perteneciente a la familia de las Tec, que son expresadas en diferentes tipos de células mieliodes.
Kinasa IP3: molécula de señalización citoplasmática, mediada por la fosfolipasa C (PLCg1) y que promueve la entrada de calcio a la célula.
Lyn: kinasas de la familia SRC
MIRR: receptores inmunes reconocedores de cadenas múltiples.
PIP3: fosfolípido que es activado posterior a la unión del antígeno y es importante para la activación del linfocito.
PLCg: enzima que cataliza la hidrólisis del fosfolípido de membrana PIP3, para generar dos moléculas de señalización que son IP3 y diacilglicerol (DAG).
Receptor Fc: receptor específico de superficie celular para la región constante carboxiterminal de la inmunoglobulina. Estos receptores son cadenas de multiproteínas que incluyen componentes de señalización y de unión a inmunoglobulinas.
Receptor Fc g: es un receptor específico de superficie, que se une a la porción carboxiterminal de la inmunoglobulina G. Existen varias clases de receptores Fc g, dentro de los cuales se incluyen los de alta afinidad (FcgRI), que intervienen en la fagocitosis en macrófagos y neutrófilos y los de baja afinidad que activan células como las NK (FcgRIIIB), o inhiben señales de transducción celular (FcgRIIB).
SH2: estructura con dominios tridimensionales con aproximadamente 100 aminoácidos, presente en muchas moléculas de señalización, que permiten uniones no covalentes con otras proteínas por unión de fosfotirosinas.
SHIP: fosfatasa que contiene inositol en su estructura y promueve la defosforilación.
SHP1 y SHP2: fosfatasas que contienen dominios SH2, encargadas de defosforilar moléculas que han sido fosforiladas previamente.
SRC: familia de proteínas con actividad kinasa, que producen la fosforilación de ITAM.
Syk: familia de moléculas que se unen a ITAM, en forma de anclaje molecular, cuando éste ya está fosforilado.
Tec: familia de kinasas que contienen dominios PH, para interactuar con PIP3.
Referencias
1. Fridman WH, Bonnerot C, DaeÈron M, Amigorena S, Teillaud JL, SauteÁs C. Structural bases of Fcg receptor functions. Immunol Rev 1992; 125: 49-76. [ Links ]
2. Berken A, Benacerraf B. Properties of antibodies cytophilic for macrophages. J Exp Med 1966; 3:119-144. [ Links ]
3. Ravetch JV, Bolland S. IgG Fc receptors. Annu Rev Immunol 2001; 19: 275-290. [ Links ]
4. Heyman B. Feedback regulation by IgG antibodies. Immunol Lett 2003; 88:157-161. [ Links ]
5. Van den Herik-Oudijk IE, Capel PJ, Van der Bruggen T, Van de Winkel JG. Identification of signalling motifs within human FcgRIIa and FcgRIIb isoforms. Blood 1995; 85:2202-2211. [ Links ]
6. Pearse RN, Kawabe T, Balland S, Guinamard R, Kurosaki T, Ravetch JV. SHIP recruitment attenuates FcgRIIb-induced B-cell apoptosis. Immunity 1999; 10:753-760. [ Links ]
7. Dijstelbloem H, Van de Winkel J, Kallenberg. Inflammation in autoimmunity receptors for IgG revisited. Trends Immunol 2001; 22: 510-516. [ Links ]
8. Takai T. Roles of Fc receptors in autoimmunity. Nat Rev Immunol 2002; 2:580-592. [ Links ]
9. Amigorena S, Bonnerot C. Fc receptor signalling and trafficking: a connection for antigen processing. Immunol Rev 1999; 172:279-284. [ Links ]
10. Wallace KH, Rees AR. Studies on the immunoglobulin-G Fc-fragment receptor from neonatal rat small intestine. Biochem J 1980; 188:9-16. [ Links ]
11. Christianson GJ, Brooks W, Vekasi S, Manolfi EA, Niles J, Roopenian SL, et al. Beta 2-microglobulin-deficient mice are protected from hypergammaglobulinemia and have defective antibody responses because of increased IgG catabolism. J Immunol 1997; 159:47814792. [ Links ]
12. Ghetie V and Sally Ward E. Multiple roles for the major histocompatibility complex class I-related receptor FcRn. Ann Rev Immunol 2000; 18:739-766. [ Links ]
13. Ravetch JV, Kinet JP. Fc receptors. Ann Rev Immunol 1991; 9:457-492. [ Links ]
14. Luanfeng P, Peng P. Signalling transduction by IgG receptors. Chinese Medical Journal 2003; 116:487-494. [ Links ]
15. Durden DL, Kim HM, Calore B, Liu Y. The Fc gamma RI receptor signals through the activation of hck and MAP kinase. J Immunol 1995; 154: 4039-4047. [ Links ]
16. Ferguson KM, Lemmon MA, Schlessinger J, Sigler PB. Structure of the high affinity complex of inositol trisphosphate with a phospholipase C pleckstrin homology domain. Cell 1995; 83:1037-1046. [ Links ]
17. Kawakami Y, Yao L, Han W, Kawakami T. Tec family protein-tyrosine kinases and pleckstrin homology domains in mast cells. Immunol Lett 1996; 54:113-117. [ Links ]
18. Wang D, Feng J, Wen R, Marine JC, Sangster MY, Parganas E, et al. Phospholipase C gamma 2 is essential in the functions of B cell and several Fc receptors. Immunity 2000; 13:2535. [ Links ]
19. Hulett MD, Hogarth PM. Molecular basis of Fc receptor function. Adv Immunol 1994; 57:1-27. [ Links ]
20. Diegel ML, Rankin BM, Bolen JB, Dobois PM, Kiener PA. Crosslinking of Fcg receptor to surface immunoglobulin on B cells provides an inhibitory signal that closes the plasma membrane calcium channel. J Biol Chem 1994; 269:11409-11416. [ Links ]
21. Muta T, Kurosaki T, Misulokin Z, Sanchez M, Nussenzweig MC, Ravetch JV. A 13-amino acid motif in the cytoplasmic domain of FcgRIIB modulates B cell receptor signalling. Nature 1994; 368:70-73. [ Links ]
22. Ono M, Bolland S, Tempst P, Ravetch JV. Role of the inositol phosphatase SHIP in negative regulation of the immune system by the receptor FcgRIIB. Nature 1996; 383:263-266. [ Links ]
23. Bolland S, Pearse RN,Kurosaki T, Ravetch JV. SHIP modulates immune receptor responses by regulating membrane association of Btk. Immunity 1998; 8:509-516. [ Links ]
24. Su K, Wu J, Edberg JC, McKenzie SE, Kimberly RP. Genomic organization of classical human low-affinity Fcg receptors genes. Genes Immun 2002 3(suppl 1) S51-S56. [ Links ]
25. Warmerdam PA, Nabben NM, van de Graaf SA, van de Winkel JG, Capel PJ. The human low affinity immunoglobulin G Fc receptor IIC gene is a result of an unequal crossover event. J Biol Chem 1993; 268: 7346-7349. [ Links ]
26. Warmerdam PA, Van de Winkel JG, Vlug A, Westerdaal NA, Capel PJ. A single amino acid in the second Ig-like domain of the human Fc gamma receptor II is critical for human IgG2 binding. J Immunol 1991; 147: 1338-1343. [ Links ]
27. Clark M, Stuart S, Kimberly RP. A single aminoacid distinguishes the high-responder from the low-responder form of Fc receptor II on human monocytes. Eur J Immunol 1991; 21: 1911-1916. [ Links ]
28. Kyogoku C, Dijstelbloem HM, Tsuchiya N, Hatta Y, Kato H, Yamaguchi A, et al. Fcg receptor gene polymorphisms in Japanese patients with systemic lupus erythematosus. Contribution of FCGR2B to genetic susceptibility. Arthritis Rheum 2002; 46: 12421254. [ Links ]
29. Wu J, Edberg JC, Redecha PB, Bansal PB, Bansal V, Guyre PM, et al. A novel polymorphism of FcgammaRIIIa (CD16) alters receptor function and predisposes to autoimmune disease. J Clin Invest 1997; 100: 1059-1070. [ Links ]
30. Huizinga TW, Kleijer M, Tetteroo PA, von Roos D, dem Borne AE. Biallelic neutrophil NA-antigen system is associated with a polymorphism on the phospho-inositol-linked Fc gamma receptor III (CD16). Blood 1990; 75: 213-217. [ Links ]
31. Salmon JE, Millard SS, Brogle NL, Kimberly RP. Fc gamma receptor IIIb enhances Fc gamma receptor IIa function in an oxidant dependent and allele-sensitive manner. J Clin Invest 1995; 95: 2877-2885. [ Links ]
32. Rose NR, Bona C. Defining criteria for autoimmune diseases (Witebsky's postulates revisited). Immunol Today 1993; 14: 426-430. [ Links ]
33. Takai T, Li M, Sylvestre D, Clynes R, Ravetch JV. FcR _-chain deletion results in pleiotrophic effector-cell defects. Cell 1994; 76: 519529. [ Links ]
34. Bolland S, Ravetch JV. Spontaneous autoimmune disease in FcgRIIB-deficient mice results from strain-specific epistasis. Immunity 2000; 13, 277285. [ Links ]
35. Van der Pol WL, van de Winkel JG. IgG receptor polymorphisms: risk factors for disease. Immunogenetics 1998; 48: 22232. [ Links ]
36. Karassa FB, Trikalinos TA, Ioannidis JP. Role of the Fcgamma receptor IIa polymorphism in susceptibility to systemic lupus erythematosus and lupus nephritis: a meta-analysis. Arthritis Rheum 2002; 46: 15631571. [ Links ]
37. Dijstelbloem HM, Scheepers RH, Oost WW, Stegeman CA, van der pol WL, Sluiter WJ, et al. Fc gamma receptor polymorphisms in Wegener's granulomatosis: risk factors for disease relapse. Arthritis Rheum 1999; 42: 18231827. [ Links ]
38. Morgan AW, Griffiths B, Ponchel F, Montague BM, Ali M, Gardner PP, et al. Fcgamma receptor type IIIA is associated with rheumatoid arthritis in two distinct ethnic groups. Arthritis Rheum 2000; 43:23282334. [ Links ]
39. Nieto A, Caliz R, Pascual M, Mataran L, Martin J. Involvement of Fcgamma receptor IIIA genotypes in susceptibility to rheumatoid arthritis. Arthritis Rheum 2000; 43: 735739. [ Links ]
40. Van Sorge NM, van der Pol WL, van de Winkel JG. FcgammaR polymorphisms: Implications for function, disease susceptibility and immunotherapy. Tissue Antigens 2003; 61:189-202. [ Links ]
41. Van der Pol WL, Jansen MD, Kuks TB, debates M, Leppers-van de Straat FG, Wokke JH, et al. Association of the Fc gamma receptor IIA-R/R131 genotype with myasthenia gravis in Dutch patients. J Neuroimmunol 2003; 144: 143-147. [ Links ]
42. Williams Y, Lynch S, McCann S, Smith O, Feighery C, Whelan A. Correlation of platelet Fc gammaRIIA polymorphism in refractory idiopathic (immune) thrombocytopenic purpura. Br J Haematol 1998; 101: 779-782. [ Links ]
43. Fujimoto TT, Inoue M, Shimomura T, Fujimura K.. Involvement of Fc gamma receptor polymorphism in the therapeutic response of idiopathic thrombocytopenic purpura. Br J Haematol 2001; 115: 125-130. [ Links ]
44. Myhr KM, Raknes G, Nyland H, Vedeler C. Immunoglobulin G Fc-receptor (FcgammaR) IIA and IIIB polymorphisms related to disability in MS. Neurology 1999; 52: 1771-1776. [ Links ]
45. Van der Pol WL, van den Berg LH, Scheepers RH, van der Bom JG, van Doorn PA, van Koningsveld R, et al. IgG receptor IIa alleles determine susceptibility and severity of Guillain-Barre syndrome. Neurology 2000. 54:1661-1665. [ Links ]
46. Breij EC, van der Pol WL, van Winsen L, Jansen MD, Dijkstra CD, van de Winkel JG, et al. No association of Fc gamma RIIa, Fc gamma RIIIa and Fc gamma RIIIb polymorphisms with MS. J Neuroimmunol 2003; 140: 210-215. [ Links ]
47. Kyogoku C, Tsuchiya N, Matsuta K, Tokunaga K. Studies on the association of Fc gamma receptor IIA, IIB, IIIA and IIIB polymorphisms with rheumatoid arthritis in the Japanese: evidence for a genetic interaction between HLA-DRB1 and FCGR3A. Genes Immun 2002; 3:488-493. [ Links ]
48. Milicic A, Misra R, Agrawal S, Aggarwal A, Brown MA, Wordsworth BP. The F158V polymorphism in FcgammaRIIIA shows disparate associations with rheumatoid arthritis in two genetically distinct populations. Ann Rheum Dis 2003; 62: 1021-1023. [ Links ]
49. Pawlik A, Ostanek L, Brzosko I, Brzosko M, Fabrycy IF, Florczak M, et al. Fc gamma RIIa polymorphism in patients with rheumatoid arthritis. Clin Exp Rheumatol 2002; 20: 841-844. [ Links ]
50. Brun JG, Madland TM, Vedeler CA. Immunoglobulin G Fc-receptor (FcgammaR) IIA, IIIA, and IIIB polymorphisms related to disease severity in rheumatoid arthritis. J Rheumatol 2002; 29:1135-1140. [ Links ]
51. Radstake TR, Petit E, Pierlot C, van de Putte LB, Cornelis F, Barrera P. Role of Fcgamma receptors IIA, IIIA, and IIIB in susceptibility to rheumatoid arthritis. J Rheumatol 2003; 30: 926-933. [ Links ]
52. Magnusson V, Zunec R, Odeberg J, Sturfelt G, Truedsson L, Gunnarsson I, et al. Polymorphisms of the Fc gamma receptor type IIB gene are not associated with systemic lupus erythematosus in the Swedish population. Arthritis Rheum 2004; 50:1348-1350. [ Links ]
53. Lee HS, Chung YH, Kim TG, Kim TH, Jun JB, Jung S, et al. Independent association of HLA-DR and Fc gamma receptor polymorphisms in Korean patients with systemic lupus erythematosus. Rheumatology 2003; 12:1501-1507. [ Links ]
54. Kyogoku C, Tsuchiya N, Wu H, Tsao BP, Tokunaga K. Association of Fcg _receptor IIA, but not IIB and IIIA, polymorphisms with systemic lupus erythematosus: a family-based association study in Caucasians. Arthritis Rheum 2004; 50: 671-673. [ Links ]
55. Siriboonrit U, Tsuchiya N, Sirikong M, Kyogoku C, Bejrachandra S, Suthipinittharm P, et al. Association of Fcg_ receptor IIb and IIIb polymorphisms with susceptibility to systemic lupus erythematosus in Thais. Tissue Antigens 2003; 61: 374-383. [ Links ]
56. Khoa PD, Sugiyama T, Yokochi T. Fc gamma receptor II polymorphisms in Vietnamese patients with systemic lupus erythematosus. Lupus 2003; 12:704-706. [ Links ]
57. Gonzalez-Escribano MF, Aguilar F, Sanchez-Roman J, Nunez-Roldan A. FcgRIIA, FcgRIIIA and FcgRIIIB polymorphisms in Spanish patients with systemic lupus erythematosus. Eur J Immunogenet 2002; 29: 301-306. [ Links ]
58. Manger K, Repp R, Jansen M, Geisselbrecht M, Wassmuth R, Westerdaal NC, et al. Fcg receptor IIa, IIIa, and IIIb polymorphisms in German patients with systemic lupus erythematosus: association with clinical symptoms. Ann Rheum Dis 2002; 61: 786-792. [ Links ]
59. Carrion F, Figueroa F, Martinez ME, Massardo L, Perez T, Foster C, et al. Polymorphisms of FcgIIa receptors in Chilean patients with systemic lupus erythematosus. Rev Med Chile 2003; 131: 11-18. [ Links ]
60. Carcao MD, Blanchette VS, Wakefield CD, Stephens D, Ellis J, Matheson K, et al. Fcgamma receptor IIa and IIIa polymorphisms in childhood immune thrombocytopenic purpura. Br J Haematol 2003; 120: 135-141. [ Links ]
61. Vedeler CA, Myhr KM, Nyland H. Fc receptors for immunoglobulin G: a role in the pathogenesis of Guillain-Barre syndrome and multiple sclerosis. J Neuroimmunol 2001; 118:187-193. [ Links ]
62. Rigby LJ, Epa VC, Mackay GA, Hulett MD, Sutton BJ, Gould HJ, et al. Domain one of the high affinity IgE receptor, Fc epsilon RI, regulates binding IgE through its interface with domain two. J Biol Chem 2000; 275: 9664-9672. [ Links ]
63. Samuelsson A, Towers TL, Ravetch JV. Anti-inflammatory activity of IVIG mediated through the inhibitory Fc receptor. Science 2001; 291: 484-486. [ Links ]
64. Kinet JP. Atopic allergy and other hypersensitivities. Curr Opin Immunol 1999; 11: 603-605. [ Links ]
65. Zusman T, Gohar O, Eliassi Y, Avivi Y, Lisanki E, Sautes C. The murine Fc-gamma (Fc gamma) receptor type II B1 is a tumorigenicity-enhancing factor in polyoma-virus-transformed 3T3 cells. Int J Cancer 1996; 65: 221-229. [ Links ]
66. Choudhry VP, Kashyap R, Ahlawat S, Pati HP. Vinblastine and danazol therapy in steroid resistant childhood chronic idiopathic thrombocytopenic purpura. Int J Hematol 1995; 61: 157-162. [ Links ]
67. Moxey-Mims MM, Noble B. Glomerular macrophage phagocytic activity in experimental immune complex nephritis. Kidney 1994; 45: 1326-32. [ Links ]
68. Kastner P, Franke H, Kleinert P, Gunther C, Malberg K, Lobnitz M. Fc receptor mediated immune complex clearance function of the mononuclear phagocyte system in systemic lupus erythematosus. Allerg Immunol 1990; 2:103-110. [ Links ]
69. Bjerke JR, Tigalonova M, Jensen TS, Matre R. Fc gamma-receptors in skin and serum from patients with psoriasis, before and after therapy. Acta Derm Venereol 1994; 186: 141-142. [ Links ]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}