










Services on Demand
Journal
Article
 Spanish (pdf)
Spanish (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mailIndicators
-
 Cited by SciELO
Cited by SciELO -
 Access statistics
Access statistics
Related links
-
 Cited by Google
Cited by Google -
 Similars in
SciELO
Similars in
SciELO -
 Similars in Google
Similars in Google
Share

 Permalink
PermalinkActa Agronómica
Print version ISSN 0120-2812
Acta Agron. vol.58 no.4 Palmira Oct./Dec. 2009
1,2Facultad de Ciencias Agropecuarias, Universidad Nacional de Colombia. AA 237. Palmira, Valle del Cauca, Colombia. 2Corporación Colombiana de Investigación Agropecuaria-Corpoica, Colombia. Autor para correspondencia: jemunozf@palmira.unal.edu.co, yaceniamc@yahoo.es
Se compararon las eficiencias de tres métodos de electroforesis en agarosa y poliacrilamida, usando la cámara pequeña de DNA Sequencing System y cámara grande OWL Sequi-Gen Sequencing Cell, en la detección del polimorfismo en 21 accesiones de naranja (Citrus sinensis) con empleo del cebador CGA. El gel de poliacrilamida dio mejor resolución de los productos amplificados vía PCR producidos por RAMs. Este permitió una mejor detección de bandas de ADN polimórficas, lo que facilitó la identificación de la variabilidad genética. La electroforesis en agarosa puede ser más conveniente en otras aplicaciones, debido al bajo costo y fácil aplicación. El estudio de diversidad genética en naranja usando microsatélites RAMs diferenció 51 accesiones en siete grupos con 0.75 de similaridad y 0.25 de heterocigosidad, lo que revela bajo polimorfismo genético. La técnica RAMs permitió agrupar las accesiones en Comunes o Blancas, Navel y Pigmentadas o Sanguinas.
Palabra clave: Citrus sinensis, electroforesis, agarosa, poliacrilamida, RAMs.
We compared the efficiency of three methods of agarose and polyacrylamide electrophoresis (using the small tank of the DNA Sequencing System and the large OWL Sequi-Gen Sequencing Cell), for the detection of polymorphism in 21 accessions of orange (Citrus sinensis), using the primer CGA. The polyacrylamide gel gave better resolution of the PCR-amplified RAM products. This method allowed better detection of polymorphic DNA bands, facilitating the identification of genetic variability. The agarose electrophoresis may be more convenient in other applications, due to its low cost and easy implementation. The study of genetic diversity in orange using RAMs separated 51 accessions into seven groups with 0.75 similarity, and 0.25 heterozygosity, revealing low genetic polymorphism. The RAMs technique grouped the accessions into Common or White, Navel and Pigmented or Sanguine.
Key words: Citrus sinensis, electrophoresis, agarose, polyacrylamide, RAMs.
El género Citrus (2n = 18) es nativo del sudeste de Asia y del archipiélago indo-malayo (Avilán et al., 1989), contiene la mandarina Citrus reticulata, limón C. medica y pomelo C. maxima los cuales son los ancestros de las especies comerciales. La apomixis facultativa es predominante en C. reticulata y ha sido determinante en la evolución de Citrus.
Los cítricos son producidos en zonas subtropicales y tropicales, sus frutas son consumidas cada día por millones de personas alrededor del mundo, la naranja Citrus sinensis L. Osbeck es la especie más representativa y reconocible de este grupo. Se originó en el sureste asiático y su híbrido característico parece provenir de un cruce entre mandarina Citrus reticulata y pomelo Citrus grandis L. Osbeck (Davies y Albrigo, 1992; Nicolosi et al., 2000).
La caracterización de los bancos de germoplasma, la variación genética y el mejoramiento en naranjas y otras especies de Citrus no han sido exitosos debido a las características relacionadas con la biología reproductiva de estas especies, por ejemplo, alta fertilidad interespecífica, reproducción apomíctica, poliembrionía, una larga fase juvenil y la escasez de marcadores de ADN polimórficos (Bretó et al., 2001; Corazza-Nunes et al., 2002).
El mejoramiento genético de los cítricos mediante métodos convencionales se encuentra limitado debido a sus características genéticas y reproductivas. Los cítricos tienen un sistema de reproducción complejo, con muchos casos de esterilidad y de ínter y autocompatibilidad, apomixis, elevada heterocigosis y la mayoría de las especies presentan un prolongado período juvenil. Además, se desconoce el modo de herencia de la mayor parte de caracteres agronómicos de interés (Grosser y Gmitter, 1990).
Las técnicas biotecnológicas tales como el cultivo de tejidos y de células y la biología molecular han ayudado a los mejoradores a sobrellevar estas dificultades. Además, la hibridación a través de la fusión de protoplastos y la transformación genética han contribuido significativamente a superar estas limitaciones (Mendes-da-Glória et al., 2000). Las técnicas moleculares han permitido el desarrollo de marcadores de ADN asociados con características de interés, así como el desarrollo del mapa de ligamiento de los cítricos, los cuales han mostrado ser útiles en la realización de una selección temprana en la progenie con los caracteres deseados en programas de mejoramiento clásico (Mendesda- Glória et al., 2000).
Los marcadores moleculares tales como el polimorfismo en la longitud de los fragmentos de restricción (RFLP), ADN polimórfico amplificado al azar (RAPD), microsatélites e isoenzimas han sido desarrollados para estudiar la diversidad genética y domesticación, establecer mapas moleculares, caracterizar variedades y ayudar en los programas de mejoramiento. Los marcadores microsatélites son simples de usar, son codominantes, tienen alto polimorfismo, y se pueden emplear para esclarecer la diversidad genética en Citrus en los niveles interespecífico, intraespecífico e intrapoblacional. La caracterización de marcadores microsatélites ha sido ya publicada en C. limon (Golein et al., 2005), C. sinensis (Ahmad et al., 2003; Novelli et al., 2006), C. limonia x P. trifoliata (Kijas et al., 1997) y C. reticulata (Koehler-Santos et al., 2003).
Los marcadores moleculares conocidos como los RAMs son útiles para medir la diversidad genética en plantas y animales, con poder discriminatorio entre familias y especies e igualmente intraespecífico (Muñoz et al., 2008), muestran la base misma de la variación de los individuos, permiten seleccionar regiones concretas dentro de la molécula de ADN para estudios determinados, el número de polimorfismos detectables es teóricamente ilimitado y permiten analizar tanto la información que se expresa como la que no lo hace (Mahuku et al., 2002). Esta metodología es factible en pequeños laboratorios en términos de equipos y facilidades de costo, no requiere el conocimiento previo de secuencias ni el uso de isótopos radioactivos (Hantula et al., 1996). Los marcadores obtenidos por los RAMs se pueden usar igualmente para estudios de poblaciones (Hantula, et al., 1996).
Se han utilizado diferentes métodos para la separación y la detección de fragmentos de ADN amplificado. La electroforesis con geles de poliacrilamida con radioactividad y tinción con plata fue el primer método usado en el análisis de los fragmentos de ADN amplificado por PCR (Cipriani et al., 1999). La electroforesis en geles de agarosa es un método utilizado como alternativa al de poliacrilamida debido a que es más barata y de fácil aplicación (Morgante et al., 2001). Recientemente, se han desarrollado nuevos métodos para el análisis de la PCR de marcadores microsatélites, basados en la utilización de un secuenciador automatizado de electroforesis capilar (Aranzana et al., 2003; Ahmad et al., 2004). Sin embargo, no se ha hecho la comparación de la eficiencia de estos métodos para la separación y análisis de los fragmentos de ADN y las implicaciones en la evaluación de la diversidad y relaciones genéticas.
El objetivo de este trabajo fue estudiar mediante la técnica RAMs la diversidad genética y comparar el polimorfismo de los métodos de electroforesis de poliacrilamida y agarosa utilizados en la caracterización molecular de naranja (Citrus sinensis).
En la caracterización molecular de la diversidad genética se utilizaron 51 accesiones de naranja C. sinensis de las 54 del banco de germoplasma de la Corporación Colombiana de Investigación Agropecuaria (Corpoica), Palmira. Para la comparación de los tres métodos de electroforesis se evaluaron 21 accesiones (Cuadro 1). La caracterización molecular se hizo en la Universidad Nacional de Colombia sede Palmira. Para la extracción de ADN se utilizó el protocolo de Dellaporta et al (1983).
Los ADN totales se visualizaron en geles de agarosa al 0.8%, teñidos con bromuro de etidio en una cámara Maxicell Primo EC-340 Electroforesis Gel System. Para determinar la concentración de ADN de cada accesión se construyó una curva de dilución con ADN del bacteriófago Lambda, de concentración inicial 20 ng/µl y se llevó a concentraciones finales de 20, 40, 60, 80 y 100 ng/µl. El ADN cuantificado se diluyó en agua tipo HPLC a un volumen total de 100 µl a 10 ng/µl y se almacenó a -20 °C.
Para el análisis RAMs se utilizaron siete cebadores polimórficos (Technologies Inc) (Bonilla et al., 2004; Mahuku et al., 2002; álvarez et al., 2003) (Cuadro 2). Para estandarizar las condiciones de cada uno de los cebadores se preparó una mezcla de reactivos en un tubo estéril de microcentrífuga (1.5 ml) para un volumen final de 25 µl (Cuadro 3).


La amplificación se llevó a cabo en un termociclador PTC 100 Programmable Termal Controller (MJ. Research, Inc). La desnaturalización inicial fue a 95 °C durante 5 min; desnaturalización a 95 °C por 30 seg, hibridación a una temperatura de 50 °C (cebador AG y CA), 55 °C (cebador CCA-TG-CT) y 58 °C (cebador GT-CGA) durante 45 seg, una extensión de 72 °C por 2 min, 37 ciclos desde la desnaturalización a extensión y por último, una extensión a 72 °C durante 7 min.
Para la comparación de los métodos de electroforesis se utilizaron 21 de las 51 accesiones de naranjas (Cuadro 1) que fueron evaluadas con el cebador CGA. Los productos amplificados fueron separados en geles de agarosa al 1.2%, a 90 voltios durante 2 h y 30 min en una cámara Maxicell Primo EC-340 Electroforesis Gel System. Posteriormente, se observaron mediante electroforesis en geles de poliacrilamida al 7% a 150 voltios por 1 h en una cámara pequeña de DNA Sequencing System. FB-SEQ-3545 de FisherBiotech. Finalmente, éstas fueron separadas en una cámara de secuenciación (OWL-Sequi-Gen Sequencing Cell) en geles de poliacrilamida al 4% (29:1 acrilamida-bisacrilamida), en condiciones denaturantes (urea 5M). A cada producto PCR se le adicionaron 10 µl de buffer denaturante (95% Formamida, 0.025% Azul de Bromofenol, 0.025% Xylene cyanol). Se cargaron 5 µl de la mezcla previamente desnaturalizadas a 95 °C durante 5 min y se corrió el gel a 120 Watts, 1600 V, durante 2 h. La tinción se realizó usando sales de plata.
Se generó una matriz binaria de ausencia (0) y presencia (1). La similitud genética entre los individuos se calculó utilizando el coeficiente de similitud de Nei y Li (1979) también conocido como DICE (1945) (Sneath y Sokal, 1973). El análisis de conglomerados se realizó con el empleo del algoritmo UPGMA y se generó un dendrograma utilizando el paquete estadístico NTSYS (Numerical Taxonomy System for personal Computer, versión 2.02 PC). Para evaluar la diversidad genética se estimó la heterocigosidad insesgada y el porcentaje de loci polimórficos utilizando el paquete estadístico TFPGA (Tools For Population Genetic Analices, versión 1.3, 1997). Se determinó el valor de F estadístico insesgado con un intervalo de confianza del 95%.
Comparación de los métodos de electroforesis
Los métodos de electroforesis geles de agarosa al 1.2% y geles de poliacrilamida fueron eficientes en la separación de los fragmentos amplificados. El primero reveló el nivel más bajo de polimorfismo en comparación con los otros dos métodos utilizados (Cuadro 4). El coeficiente de diferenciación genética (Fst) fue 0.20, con una desviación estándar (DE) de 0.04, lo que ratificó la existencia de una diferenciación genética intermedia.
Los geles de agarosa permiten una electroforesis rápida, pero con una resolución limitada por cuanto las bandas formadas tienden a ser difusas y a expandirse, debido al tamaño de los poros lo que no puede ser controlado (Westermeier, 1997). No obstante tienen la ventaja de favorecer un amplio rango de separación para fragmentos de alto peso molecular (100 pb a 50 Kpb) (Fernández-Tresguerres, 2003). Este tipo de electroforesis es preferido para la resolución de fragmentos de ADN resultantes de la amplificación vía PCR usando marcadores RAPDs (Williams et al., 1990). Sin embargo, cuando se generan varios fragmentos de ADN de tamaños similares, como ocurre a menudo en la amplificación de ADN fingerprinting, la agarosa no resulta eficiente (Caetano-Anollés, 1991).
El número de bandas obtenido por el cebador CGA en geles de poliacrilamida cámara pequeña fue de 31. El porcentaje de loci polimórficos fue 16.13%, con una heterocigosidad de 0.09 (Cuadro 4). En comparación con el gel de agarosa, en el presente trabajo se obtuvo mayor número de bandas ya que la poliacrilamida permite una mejor resolución y separación de los productos amplificados y, por tanto, mayores valores de heterocigosidad. Aunque el valor de Fst fue mayor que en el de agarosa (0.23), lo que revela una diversidad genética intermedia.
Cuando los fragmentos amplificados fueron separados en la cámara grande OWL, se obtuvo el mayor número de bandas (53), con un 74% de loci polimórficos, una heterocigosidad promedio esperada de 0.33 y un Fst de 0.09 (Cuadro 4). La poliacrilamida es un soporte empleado frecuentemente en electroforesis en gel, siendo químicamente inerte, de propiedades uniformes, y de preparación rápida y reproducible. Forma geles transparentes con estabilidad mecánica, insolubles en agua, relativamente no-iónicos, que permiten buena visualización de las bandas durante tiempo prolongado. Además tiene la ventaja de que al variar la concentración de polímeros se puede modificar de manera controlada el tamaño del poro (Campbell, 1995). Las diferencias entre los tres métodos de electroforesis son importantes, ya que métodos de mayor poder de resolución permiten detectar mayores diferencias.
Es posible utilizar el método de electroforesis de poliacrilamida para identificar variación alélica entre cultivares de naranja a una escala más fina; sin embargo, la detección es más costosa en términos de equipos y análisis, usa reactivos más tóxicos, requiere mayor tiempo y personal calificado.
En forma general, en el gel de poliacrilamida, cámara pequeña, las bandas primarias u oscuras frecuentemente coalescen con las secundarias, dando bandas tenues, especialmente aquellas de menor peso molecular. Por el contrario, las bandas en el gel poliacrilamida de cámara grande fueron más nítidas, de mayor tamaño y de grosor uniforme y las bandas primarias y secundarias fueron identificadas con relativa facilidad. Las bandas primarias presentaron buena resolución en agarosa, pero las secundarias no aparecieron claramente. El background fluorescente de bromuro de etidio redujo significativamente la claridad de los fragmentos de ADN, especialmente en las bandas tenues o secundarias.
El uso de poliacrilamida en cámara grande permite una mejor identificación de bandas polimórficas que los otros métodos usados, ya que bandas solas son resueltas en dos bandas, además, no utiliza compuestos tóxicos como el bromuro de etidio que es mutagénico. La electroforesis fue más rápida y más tolerante a temperaturas más elevadas de corrida y esto permitió el uso de voltajes más altos (100 Watt). El gel de poliacrilamida se crece durante la tinción, lo que hace que sea más fácil distinguir bandas y calcular el polimorfismo, además se pueden almacenar por largos períodos sin ninguna distorsión de los frentes de corrida. Una ventaja importante de los geles de poliacrilamida es que son químicamente inertes, transparentes y estables en un amplio rango de pH, temperatura y fuerza iónica.
Los geles de agarosa y poliacrilamida se pueden emplear en multitud de tamaños, grosores y porosidades y permiten desarrollar la electroforesis en diferentes configuraciones. La elección depende principalmente del tamaño de los fragmentos a separar.
Los geles de poliacrilamida en vertical son el método de elección más efectivo en la separación de pequeños fragmentos de ADN (5 a 500 pb). El poder de resolución es tan alto, que es posible separar fragmentos de ADN que difieren en tamaño en 1 pb, como ocurre en el caso de la secuenciación de ADN. Los principales limitantes de los geles de policiacrilamida son la neurotoxicidad de la acrilamida y la dificultad de preparación y manejo. El estudio mostró que el gel de poliacrilamida da una mejor resolución de los productos amplificados vía PCR producidos por RAMs. Este método permitió una rápida electroforesis de fragmentos de ADN y facilitó una mejor detección de bandas de ADN polimórficas, facilitando la identificación de la variabilidad genética existente en poblaciones naturales. Por tanto, el análisis de los productos amplificados de PCR por electroforesis de agarosa tiene que ser completado con la electroforesis de poliacrilamida, debido a su más bajo nivel de precisión. El método de agarosa puede ser utilizado en otros estudios debido a su bajo costo y fácil aplicación.
De acuerdo con los resultados, la técnica RAMs puede ser una herramienta útil para caracterizar la diversidad genética de diferentes accesiones dentro del género Citrus, además es una técnica que no necesita información previa, utiliza un cebador, puede ser reproducible por el tamaño del cebador, diferencia entre especies y variaciones intraespecíficas, detecta relaciones entre grupos biológicos y grupos formados por la técnica, es de bajo costo y de fácil implementación (Muñoz et al., 2008).
Estudio de diversidad genética con electroforesis en cámara de secuenciación
Los siete cebadores RAMs generaron 176 bandas; que fluctuaron entre 32 (CCA) y 55 (TG). El número de loci polimórficos varió entre 18 y 34 para los cebadores CT y CGA, respectivamente (Cuadro 5). El análisis mediante el coeficiente de Nei-Li, a un nivel de similitud de 0.75, diferenció la población en siete grupos (Figura 1). El grupo 1 estuvo conformado por naranjas comunes, Jaffa, Indian River, Cuban Queen, Salustiana, de frutos esféricos, achatados o elipsoidales, de medianos a grandes y sin ombligo, con coloraciones desde amarillo-naranja a naranja intenso. Algunas variedades tienen numerosas semillas. En el grupo 2 se localizaron la mayoría de las accesiones tipo Navel u ombligonas, sus frutos son partenocárpicos y carecen de semillas lo cual contribuye a que los individuos sean similares. El principal destino de frutos del grupo Navel es el consumo en fresco y una pequeña parte se destina a la industrialización debido a que el contenido en zumo es elevado y la presencia de limonina da lugar a un sabor amargo. Dentro del grupo 2 también se situaron algunas accesiones de naranjas comunes: Pera del Río, Lima Dulce y Enterprise, entre otras, poniendo de manifiesto la existencia de relaciones genéticas entre los tipos de naranjas evaluadas. En los grupos 3 a 6 se encontró una combinación de diferentes tipos de naranjas blancas, Valencia o comunes, lo que pone de manifiesto el continuo intercambio de materiales entre los agricultores. Las naranjas pigmentadas o tipo sanguina se diferenciaron del resto de accesiones evaluadas (grupo 7) por la presencia de antocianinas en la pulpa y a veces en la epidermis. Las naranjas sanguinas mantienen y superan los beneficios de la naranja común, ya que aportan más betacarotenos, además son interesantes en la industria ya que su sabor recuerda ligeramente al de las cerezas o las frambuesas.
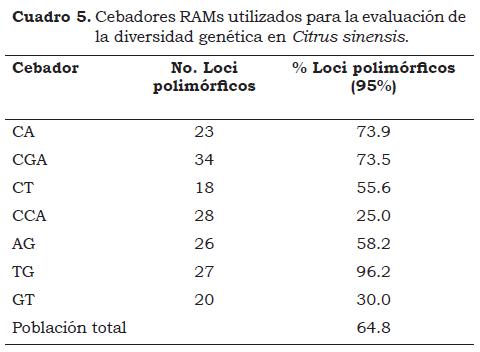
La heterocigosidad promedio (He) para la población total fue 0.25 y reveló un bajo polimorfismo genético. El cebador TG hizo el mayor aporte a la variación genética (Cuadro 6). Novelli et al (2006) desarrollaron y caracterizaron marcadores moleculares microsatélites polimórficos para naranja dulce C. sinensis L, encontrando valores de He en un rango comprendido entre 0.49 y 0.57 con un promedio de 0.50, una heterocigosidad observada (Ho) entre 0.88 y 1.00 y una Ho promedio de 0.99, que son valores altos en comparación con los encontrados en el presente estudio.

Tapia et al. (2005) evaluaron 63 cultivares de mandarina (Citrus spp.) usando marcadores morfológicos y AFLP (Amplified Fragment Length Polymorphism) para 20 caracteres cuantitativos y 10 cualitativos de hojas, flores y frutos. Las mejores combinaciones de iniciadores AFLP fueron la Mse + CAG más Eco + ACA, y Mse + CAA más Eco + AGG, dando un total de 109 bandas con un 86% de polimorfismo. Tanto los marcadores morfológicos como los moleculares mostraron un alto grado de variación entre los individuos analizados, lo que indica que ésta es una fuente importante de variabilidad para programas de mejoramiento genético. Aunque la comparación de los resultados morfológicos y moleculares, usando la prueba de Mantel, no mostró una correlación significativa (r = 0.31), ambas técnicas parecen ser complementarias para la caracterización de mandarinas.
Golein et al (2005) reportaron el aislamiento y caracterización de siete loci microsatélites polimórficos en Citrus. Estos marcadores produjeron entre 4 y 9 alelos por locus (con un promedio de 6.14) en 32 cultivares de C. limon evaluados. Los valores de heterocigosidad promedio observada estuvieron comprendidos entre 0.43 y 0.72. Los niveles de polimorfismo encontrados en este estudio sugieren que estos loci microsatélites pueden ser una herramienta importante para estudios genéticos en Citrus.
En el presente estudio el coeficiente de diferenciación genética (Fst) obtenido al evaluar las 51 accesiones de naranja con los siete microsatélites RAMs fue de 0.07, con una desviación estándar de 0.01 (Cuadro 7). Según Wrigth (1978) valores comprendidos entre 0 y 0.05 muestran poca diferenciación genética, de 0.05 a 0.15 moderada, y mayores de 0.25 una gran diferenciación. Teniendo en cuenta lo anterior, el valor de Fst encontrado en este estudio muestra que existe una diferenciación genética intermedia.

La baja diferenciación genética encontrada en C. sinensis, al igual que en los géneros Fortunella y Poncirus, se debe a que cada una de sus numerosas formas cultivadas ha sido propagada a partir de un progenitor único por vía asexual a través de injertos, esquejes, acodos y embrionía nucelar, lo que asegura el mantenimiento de la misma constitución genética en la descendencia que forma una variedad hortícola (cultivar) o clonal (Webber, 1943).
La selección de las formas cultivadas ha buscado el mejoramiento de los caracteres útiles para el hombre, como los relacionados con las cualidades del fruto –sabor, contenido del zumo, azúcar y ácido, textura de la pulpa, grado de aspermia, color de la epidermis, forma y talla–, el comportamiento y las características del árbol –vigor, productividad, y forma no espinosa–. Por consiguiente, existe una tendencia a la uniformidad en el proceso de domesticación de cada especie y, por esta razón, entre ciertas variedades hay tantas semejanzas en lo que se refiere a los caracteres más visibles, que resulta difícil diferenciarlos (Webber, 1943).
Se sabe que las naranjas dulces tienen una estrecha base genética y que la variabilidad puede ser producida por diversos factores tales como hibridación, mutación y tipo de reproducción. La baja diversidad intraespecífica contrasta con la alta variabilidad en cuanto a características de importancia agronómica tales como período de maduración, tamaño y color de los frutos (Webber, 1943).
- La electroforesis con el uso de geles de poliacrilamida en la cámara OWL Sequi- Gen Sequencing Cell, permitió una mejor resolución de los productos amplificados ayudando así a la detección del polimorfismos entre las 21 accesiones de naranja C. sinensis evaluadas.
- Los valores de similaridad de Nei-Li y de heterocigosidad revelaron un bajo polimorfismo genético, que puede ser debido a la propagación vegetativa de la especie, la cual tiende a perpetuar genotipos similares a través de varias generaciones.
- La técnica RAMs permitió agrupar las diferentes accesiones de naranjas según los tipos reportados (Comunes o Blancas, Navel y Pigmentadas o Sanguinas), lo que indica que esta es una herramienta útil para evaluar la diversidad genética en las naranjas evaluadas y que podría ser útil para otras especies de Citrus.
Los autores expresan sus agradecimientos al grupo de Diversidad Biológica de la Universidad Nacional de Colombia sede Palmira, a la División de Investigación DIPAL de la Universidad Nacional de Colombia, Sede Palmira y a la Corporación de Investigación Agropecuaria (Corpoica)-Palmira.
Ahmad, R; Struss, D; y Southwick, S. M. 2003. Development and characterization of microsatellite markers in Citrus. J.Amer. Societ.Hort Sci, 128:584 - 590. [ Links ]
Ahmad, R; Potter, D. and Southwick, S.M. 2004. Identification and characterization of plum and plumcot cultivars by microsatellite markers. J. Hort. Sci. Biotechnol. 79 (1):164-169. [ Links ]
Alvarez, E; Mejia, J.F; and Valle, T.L. 2003. Molecular and pathogenicity characterization of Sphaceloma manihoticola isolates from south-central Brazil. Plant Dis. 87:1322 - 1328. [ Links ]
Aranzana, M.J; Cosson, P; Dirlewanger, E; Ascasibar, J; Cipriani, G; Arus, P; Testolin, A; King, G.J. y Lezzoni, A. F. 2003. A set of simple-sequence repeat (SSR) markers covering the Prunus genome. Theor. Appl. Genet. 106(5):819-825. [ Links ]
Avilán, L; F. Leal, y D. Bautista. 1989. Manual de fruticultura. Editorial América, C.A. Caracas, Venezuela. p 1316-1317. [ Links ]
Bonilla, M; Espinosa, K; Muñoz, J.E y Vásquez, H. 2004. Colección, caracterización fenotípica y molecular de poblaciones de uchuva Physalis peruviana L. Congreso Colombiano de Botánica: Botánica, Diversidad y Cultura. 2, Popayán, Noviembre de 2004. p. 350 - 351. [ Links ]
Bretó, M. P; Ruiz, C; Pina, J. A; y Asíns, M. J. 2001. The diversification of Citrus clementina Hort. ex Tan., a vegetatively propagated crop species. Mol. Phylogenet. Evol 21:285 - 293. [ Links ]
Caetano-Anollés, G; Bassam, B. J.; y Gresshoff, P. m. 1991. DNA amplification fingerprinting using very short arbitrary oligonucleotide primers. Biotechn. 9: 53 - 557. [ Links ]
Campbell, M. K. 1995. Biochemistry. Second edition; Saunders College Publishing: Orlando, FL, 1995; p. 522. [ Links ]
Cipriani, G; Lot, G; Huang, W.G; Marrazzo, M.T; Peterlunge, E; y Testolin, R. 1999. AC/GT and AG/CT microsatellite repeats in peach (Prunus persica (L.) Basch): isolation, characterization and cross-species amplification in Prunas. Theor. Appl. Genet. 99(1-2):65-72. [ Links ]
Corazza-Nunes, M. J; Machado, M. A; Nunes, W. M.; Cristofani, M. y Targon M. L. 2002. Assessment of genetic variability in grapefruits (C. paradisi Macf.) and pummelos (C. maxima (Burm.) Merr.) using RAPD and SSRs markers. Euphytica 126:169 - 176. [ Links ]
Davies, F. y Albrigo, L. G. 1992. Taxonomy, cultivars and breeding. En: Gmiter Jr F. G; Grosser, J. W. y Moore G. A (eds.). Citrus. CAB International, Wallingford. p. 12 - 51. [ Links ]
Dellaporta, S. l.; Wood, J. y J. B. Hicks. 1983. A plant DNA minipreparation: Versión II. Plant Mol. Biol. Rep. 1(14):19-21. [ Links ]
Dice, L. R. 1945. Measures of the amount of ecological association between species. Ecology. 26:297 - 302. [ Links ]
Fernández.-Tresguerres, J. A. 2003. Biotecnologia aplicada a la medicina. Ediciones Díaz de Santos, Madrid. 352 p. [ Links ]
Golein, B; Koltunow, A. M; Talaie, A; Zamani, Z.; y Ebadi, A. 2005. Isolation and characterization of microsatellites loci in the lemon (Citrus limon). Mol. Ecol. Notes 5:253 - 255. [ Links ]
Grosser, J. W y Gmitter, Jr. F. G. 1990. Protoplast fusion and Citrus improvement. Plant Breeding Rev. 8:339 - 374. [ Links ]
Hantula, J; Dusabenyagasani, M; Hamelin, R. C. 1996. Random Amplified Microsatellites (RAMs) a novel method for characterizing genetic variation within fungi. Eur. J. For. Path. 26: 159-166. [ Links ]
Henríquez, N. M. 2000. Diversidad genética de Phaeoisariopsis griseola (Sacc) Ferraris, utilizando marcadores moleculares. Trabajo de grado Ing Agr. Universidad Nacional de Colombia, sede Palmira. 101 p. [ Links ]
Koehler-Santos, P.; Dornelles, A. L. y Freitas, L. B. 2003. Characterization of mandarin citrus germplasm from southern Brazil by morphological and molecular analyses. Pesqu. Agropec. Bras. 38:797 - 806. [ Links ]
Kijas, J. M.; Thomas, M. R.; Fowler, J. C. y Roose, M. L. 1997. Integration of trinucleotide microsatellites into a linkage map of Citrus. Theor. Appl. Gen. 94:701 - 706. [ Links ]
Mahuku, G. S.; Henríquez, M. A.; Muñoz, J. E y Buruchara, R. A. 2002. Molecular markers dispute the existence of the Afro- Andean group of the Bean Angular Leaf Spot pathogen, Phaeoisariopsis griseola. Phytopath. 96(6):580 - 589. [ Links ]
Mendes-Da-Glória, F. J.; Mourão Filho, F. A.; Aranha-Camargo, L. E y Mendes, B. M. 2000. Caipira sweet orange + rangpur lime: a somatic hybrid with potential for use as rootstock in the brazilian citrus industry. Gen. Mol. Biol. 23(3):661 - 665. [ Links ]
Morgante, M.; Pfeiffer, A.; Jurman, I.; Paglia, G.; y Olivieri, A. M. 2001. PCR analysis of SSR polymorphisms in plant using agarose gels. En: Karp, A; Isaac, P. G; Ingram, D.S (eds.). Molecular tools for screening biodiversity, Kluwer Academic Publ. Dordrecht, Holanda. p. 206 - 207. [ Links ]
Muñoz, J. E.; Morillo, C. A.; Morillo, C. Y. 2008. Microsatélites Amplificados al Azar (RAM) en estudios de diversidad genética vegetal. Acta Agron. 57(4):219 - 226. [ Links ]
Nei, M. y Li, W.H. 1979. Mathematical model for studying genetic variation in terms of restriction endonucleasa. Proc. Nat. Acad. Sci. 79:5267 - 5273. [ Links ]
Nicolosi, E.; Deng, Z.N.; Gentile, A.; La Malfa, S.; Continella, G.; y Tribulato, E. 2000. Citrus phylogeny and genetic origin of important species as investigated by molecular markers. Theor. Appl. Genet. 100:1155 - 1166. [ Links ]
Novelli, V.M.; Cristofani, M.; Souza, A. A. y Machado, M. A. 2006. Development and characterization of polymorphic microsatellite markers for the sweet orange (Citrus sinensis L. Osbeck). Gen. Mol. Biol. 29(1):90-96. [ Links ]
Sneath, P. H. y Sokal, R. R. 1973. Numerical taxonomy: the principles and practice of numerical classification. W.H. Freeman & Co., San Francisco. 573 p. [ Links ]
Tapia, C. E.; Espinosa; G. M.; Warburton, L. M.; Varela, S. A; y Monter, V.A. 2005. Characterization of mandarin (Citrus spp.) using morphological and AFPL markers. Interciencia: Rev. Ciencia y tecnología de América 30(11):687 - 693. [ Links ]
Webber, H. J. 1943. Plant characteristics and climatology. En: H.J. Webber y L.D. Batchelor, dirs. The Citrus Industry. Berkeley: University of California. Press. 1:41-69. [ Links ]
Westermeier, R. 1997. Electroforesis in Practice: a Guide to Methods and Applications of DNA and Protein Separations (2a. ed.), VCH Press Weinheim. [ Links ]
Williams, J. G.; Kubelik, K. J.; Livak, J. A.; Rafalski y, S. V. y Tingey. 1990. DNA polymorphisms amplified by arbitrary primers are useful as genetic markers. Nucleic Acids Res. 18:6531 - 6535. [ Links ]
Wright, S. 1978. Evolution and the genetics of populations, variability within and among natural populations, University of Chicago Press. 4:335. [ Links ]
1 Ing. Agrónoma, Ph.D. en Fitomejoramiento y Producción de Semilla.
2 Ing. Agrónoma, Ph.D. en Fitomejoramiento y Producción de Semilla.
3 Ing. Agrónoma.
4 Ing. Agrónomo, M.Sc. en Citricultura.
5 Ing. Agrónomo, M.Sc. en Genética y Mejoramiento de Plantas, Ph.D. en Horticultura.
6 Biólogo, Especialización en Microbiología.
7 Ing. Agrónoma, M.Sc. en Fitomejoramiento
8 Ing. Agrónomo, Especialización en Fruticultura Cítrico
9 Ing. Agrónomo, M.Sc. Ph.D. en Ciencias Agrarias con énfasis en Mejoramiento Vegetal.

