











 English (pdf)
English (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mail Cited by SciELO
Cited by SciELO  Cited by Google
Cited by Google  Similars in
SciELO
Similars in
SciELO  Similars in Google
Similars in Google 
 Permalink
PermalinkIntroduction
The main rootstocks used in the pear seedlings are those of the Cydonia genus, especially Cydonia oblonga (Mill.). In general, these combinations result in plants of lower vigor, with early entry into production and greater productivity. However, the reduction in vigor that occurs in the pear/quince combinations can be attributed to the graft incompatibility phenomenon (Pina et al., 2012). Graft incompatibility is the abnormal development of the graft due to anatomical, physiological, biochemical, and molecular differences, resulting in damage to the graft union, in addition to delayed vegetative growth, low productivity, and in severe cases, death of the plant (Davies et al., 2017, p. 1024; Dogra et al., 2018).
Graft incompatibility can cause major damage to commercial orchards by negatively affecting plant vegetative growth and thus reducing the potential yield (Machado et al., 2017). But there may be different levels of incompatibility depending on the pear and quince cultivars grafted. Among the main quince rootstocks used in combinations with pear trees are the ‘EMC’, highly dwarfing, and ‘Adams’, of greater vigor (Francescatto et al., 2010).
The graft incompatibility of pear/quince combinations occurs due to problems in the vascular connection of the graft, especially in the regeneration of the cortex and phloem, occurring elevated percentage of necrotic area in the graft union (Balbi et al., 2019). However, this lack of vascular continuity between pear and quince is attributed, in many cases, to the release of cyanide at the graft interface, a process known as cyanogenesis, which occurs through the hydrolysis of CGs (Gur et al., 1968; Moore, 1986; Nocito et al., 2010).
In pear/quince combinations, the CG prunasin, which occurs in the quince and not in the pear, rises from the rootstock to the cultivar, where it is hydrolyzed by β-glycosidase, causing the release of cyanide at the graft interface (Gur et al., 1968). Thus, the levels of CGs in scion and rootstock can serve as indicators of compatibility (Gur & Blum, 1973; Pereira et al., 2018).
In general, cyanogenic species also have the ability to metabolize cyanide, preventing cell damage. The cyanide detoxification process consists of two main routes. The first route involves the formation of β-cyanoalanine from cyanide and cysteine, a reaction catalyzed by β-cyanoalanine synthase. β-cyanoalanine is later converted to asparagine (Miller & Conn, 1980). The second route converts cyanide to thiocyanate and is catalyzed by rhodanase (Bordo & Bork, 2002). The most common route in plants is β-cyanoalanine (Zagrobelny et al., 2004). In this context, it can be suggested that the degree of graft compatibility or incompatibility depends both on the levels of CGs and on their ability to metabolize the released cyanide.
The objective of this work was to evaluate the growth and development of the plants, symptoms of graft incompatibility, and the relationship of these parameters with the concentration of CGs amygdalin and prunasin in the cultivar and in the rootstock of pear grafts, establishing a compatibility indicator. The tested hypothesis is that the concentrations of CG in the crown and in the rootstock are directly related to the degree of graft compatibility and to the growth of the plant.
Material and methods
The work was carried out in two private orchards located in the Arroio do Padre, Rio Grande do Sul, Brazil (Coordinates: 31° 26’ 35.51’’ S, 52° 25’ 22.10’’ O and altitude: 251 m). As the orchards had different combinations and ages, they were evaluated as different experiments.
Plant material
Experiment 1 evaluated combinations between ‘Cascatense’ (P. communis L.) cultivar and P. Calleryana (L.) and ‘EMC’ (Cydonia oblonga Mill.) rootstocks. The orchard was implanted in the winter of 2009, with spacing of 1.5 m between plants and 5.0 between lines. Experiment 2 evaluated the P. communis (L.) cultivars, ‘Packham’s Triumph’, ‘Santa Maria’, and ‘Rocha’, grafted on the ‘Adams’ quince rootstock (C. oblonga Mill.). This orchard was implanted in the winter of 2011, with a spacing of 1.0 m between plants and 5.0 between lines.
Orchard management
The orchards were conducted on terrace of 0.5 m high and 1.0 m base, with drip irrigation during the summer. The lines and inter-lines were maintained with plant cover (Avena strigosa), managed with mowing and herbicide applications.
Fertilization was carried out using urea (45 % N), triple super phosphate (42 % P2O5), and potassium chloride (58 % K2O). With 50 % of N applied in post-harvest, between March 15 and April 15, and the remainder, during the bud swelling period; P and K were applied during the winter dormancy period.
The region has insufficient cold for the evaluated cultivars, with a historical average ranging from 322 to 496 chilling hours, with dormancy breaking with hydrogenated cyanamide and mineral oil being indicated.
Growth, production and compatibility assessments
Vegetative growth assessments were carried out in October 2014, and consisted of measuring trunk diameter, plant height and vigor. The trunk diameter (cm) was evaluated 5 cm above (scion) and below (rootstock) of the grafting point, using a digital caliper. The plant height (cm) was determined from the grafting point to the apex of the plant, with a graduated measuring tape. Yield (kg ha-1) was obtained by harvesting fruits of all plants in each plot. The plant vigor was evaluated by classifying the combinations into five classes: ‘Very low’, ‘Low’, ‘Medium’, ‘High’, and ‘Very high’, being carried out by three evaluators.
In February, the branches length (cm) of the year was also evaluated by measuring 15 branches per plant. The defoliation was estimated by classifying the combinations in to four classes:0 (without defoliation),1 (defoliation of medium intensity),2 (intense defoliation), and (complete defoliation). Three evaluators performed operations.
Characterization of graft compatibility by growth parameters, visual symptoms, and biochemical responses as for symptoms, and combinations were classified (subjectively by three evaluators) according to the intensity of the symptoms in: ‘Absent’, ‘Moderate’, and ‘Severe’. Taking into account in this classification the presence of swelling in the graft region, diameters of the cultivar and rootstock, the presence of lumps in the region close to the graft union, and density of shoot emission from the rootstock.
Determination of cyanogenic glycosides (CGs)
CGs concentration was determined in the bark tissues from the scion (5 cm above graft union) and rootstock (5 cm below graft union) in the spring (October 2014) and summer (February 2015). The bark samples were immediately frozen in liquid nitrogen, stored at -80 °C and subsequently lyophilized.
The extraction was carried in 500 mg of lyophilized sample, with 8 ml of methanol and 2.000 mg of activated carbon. This mix remained under stirring for 16 h at room temperature. Then, a centrifugation was performed at 5600 g for 20 min and the supernatant was vacuum filtered through a 0.1 µm nylon filter. 20 µL of the filtered solution were subjected to chromatographic analysis. An Inertsil ODS-3 analytical column (150 x 4.6 mm, 4 µm) was used at 25 °C during the analysis. The mobile phase was water-methanol (60:40; v/v) with a flow of 1.3 mL min-1 and UV detection at a wavelength of 254 nm. Were used prunasin and amygdalin standards (Sigma-Aldrich), activated carbon (Sigma-Aldrich), and methanol (Vetec), specifics for HPLC (High Performance Liquid Chromatography). The water used was obtained employing the Milli-Q system purification (Millipore) and filtered through 0.1 mm nylon filter.
Chromatographic separation of the amygdalin and prunasin was successfully performed, with four peaks: Peak 1: water (1.16 min), Peak 2: methanol (1.38 min), Peak 3: amygdalin (2.44 min), and Peak 4: prunasina (3.44 min). The results were expressed in mg g-1 of the CGs on dry basis.
Statistical analysis
The experiments were analyzed independently. The experimental design was in randomized blocks with four replications and each experimental unit consisted of five plants. The results were subjected to variance analysis, and variables with significant differences were compared using Tukey test, at 5 % probability of error.
Results and discussion
Experiment 1
In Experiment 1, the Cascatense/P. calleryana showed a larger trunk diameter in the scion and rootstock, as well as a higher plant height and yield (Figures 1a, 1b, 1c). The high vigor of the Cascatense/P. calleryana combination was already an expected result, as P. calleryana is considered a vigorous rootstock, that is, it provides intense vegetative growth to the cultivar (Francescatto et al., 2010; Melo et al., 2017).
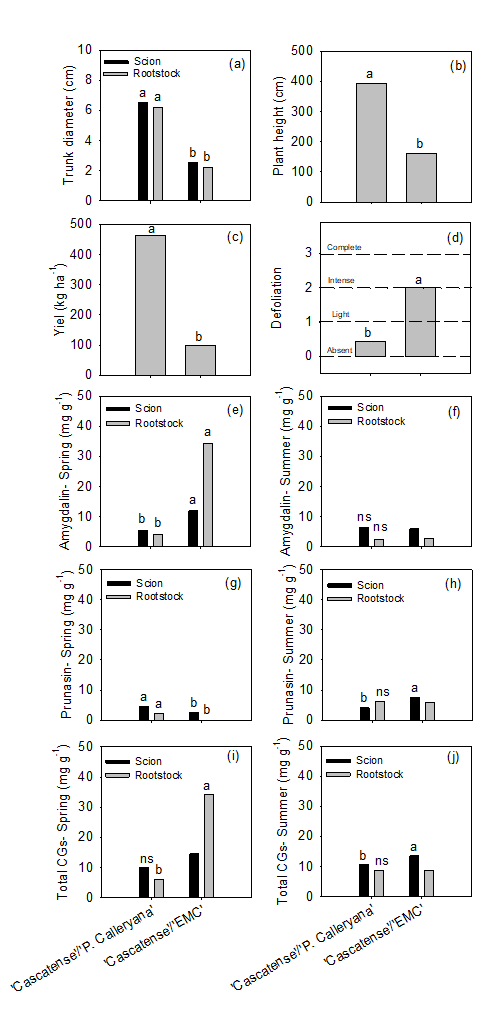
Figure 1. Growth, yield and biochemical assessments. *Different lowercaseletters differ by Tukey’s test at 5 % probability of error.
On the other hand, the ‘EMC’ rootstock is used to reduce the vigor of the cultivar canopy and increase production (Maas, 2008). However, it was found in the present study that in addition to vigor reduction, there was also a decrease in yield, suggesting that graft incompatibility between ‘Cascatense’ and ‘EMC’ occurs at a very high level. This fact can be observed both in the vigor and compatibility assessment (Figure 2). Cascatense/EMC combination was classified as ‘Very low’ vigor and with graft incompatibility symptoms ‘Severe’ (Figure 2). The incompatibility observed between ‘Cascatense’ and ‘EMC’ also explains the high defoliation rate of this combination, classified as ‘Intense’ (Figure 1d), a phenomenon that also compromises the productive capacity of the next harvest. The incompatibility between ‘Cascatense’ and ‘EMC’ has already been noted by Giacobbo et al. (2007), who even found the same symptom of marked hypertrophy at the graft union point, in addition to overgrowth and reduced diameter above and below the grafting point. The incompatibility in pear/quince combinations is of the ‘localized’ type, which manifests itself in the form of structural disorganization in the graft union region (Ermel et al., 1999), mainly due to problems continuity in cortex and phloem (Balbi et al., 2019). In advanced stages, this type of incompatibility can cause the depletion of the root system, which occurs due to poor translocation at the point of union (Errea et al., 1994; Zarrouk et al., 2006).

Figure 2. Vigor, graft incompatibility symptoms, and biochemical assessments. *Vigor classes: ‘Very low’, ‘Low’, ‘Medium’, ‘High’, and ‘Very high’. Grafting incompatibility visual symptoms: ‘Absent’, ‘Light’, ‘Moderate’, and ‘Severe’. (ns) does not differ and (**) differs by Tukey's test with 1 % probability of error.
In the spring, the levels of amygdalin were higher both in the cultivar and in the rootstock from Cascatense/EMC (Figure 1e), while in the summer there were no differences (Figure 1f). About prunasin, it was observed that in spring the concentrations were higher in the scion and rootstock from Cascatense/P. calleryana (Figure 1g). In summer, the prunasin content was higher in the cultivar ‘Cascatense’ grafted on ‘EMC’ (Figure 1h).
In general, prunasin concentrations in the cultivar and in the rootstock from two combinations were quite low compared to those of amygdalin, indicating a greater role of amygdalin in the incompatibility process. However, according to Gur et al. (1968), the CG responsible for the incompatibility of grafting between pear and quince should be prunasin, which according to the same authors, occurs in quince and not in pear. But, in the present study, in addition to having determined that both CGs are present in the pear and quince tree, the CG that seems to have the greatest relationship with the incompatibility process is amygdalin.
However, it is important to note that amygdalin is a prunasin precursor (Conn, 1980), so depending on the season, the age of the plants, the genotypes studied, among others, it is possible that one or other is determined in greater concentration.
The amygdalin and the β-glycosidase enzyme are compartmentalized in the tissues from cyanogenic plants, and the cyanogenesis process starts only when these compartments rupture, probably due to the grafting process. From that moment, the amygdalin is hydrolyzed by β-glycosidase, generating one molecule of the glucose and another one of prunasin and later, prunasin is hydrolyzed by the same enzyme, producing another molecule of glucose and an α-hydroxylitrile, which finally suffers action of α-hydroxylitrile lyase, resulting in cyanide release (Conn, 1980; Zagrobelny et al., 2004). Thus, perhaps the most appropriate is to consider the cyanogenic potential as the concentration of total CGs (amygdalin + prunasin) in the graft components.
In this sense, in the samples collected in the spring, there were no differences between the levels in the cultivar grafted on the two rootstocks (Figure 1i). However, there was a significant difference in the total content of CGs between rootstocks, with 'EMC' showing 5.6 times more than Pyrus Calleryana (Figure 1i). In summer, the total content of CG was higher in the cultivar ‘Cascatense’ grafted on ‘EMC’ (Figure 1j). However, there were no differences between the rootstocks of the two combinations (Figure 1j).
The differences between levels of total CGs from scion and rootstock of each combination were observed in the Cascatense/P. Calleryana, both in spring and summer, with no significant differences. The fact that probably contributes to its good compatibility. On the other hand, in the combination Cascatense/EMC a significant difference was found in the concentration of total CGs between scion and rootstock (Figure 2). It was found that the rootstock 'EMC' showed a concentration of total CGs 19.84 mg g-1 higher than 'Cascatense' or 2.4 fold higher. A difference that is possibly associated with the graft incompatibility observed in this combination. Pereira et al. (2018) observed in incompatible Prunus combinations, a difference of 1.9 fold.
It was also possible to observe that even in the pear/pear combination (P. communis/P. Calleryana), significant amounts of amygdalin and prunasin were found. Indicating that, differently from what was suggested by Gur et al. (1968), the pear has cyanogenic glycosides in its tissues, however, in concentrations lower than that found in quince trees.
Experiment 2
In Experiment 2, Packham’s Triumph/Adams combination showed a larger trunk diameter in the scion and rootstock, in addition to greater plant height and length of branches (Figures 3a, 3b, 3c). Regarding defoliation, the combination Santa Maria/Adams suffered ‘Intense’ defoliation, being superior to the others, which had only a defoliation level considered ‘Light’ (Figure 3d).
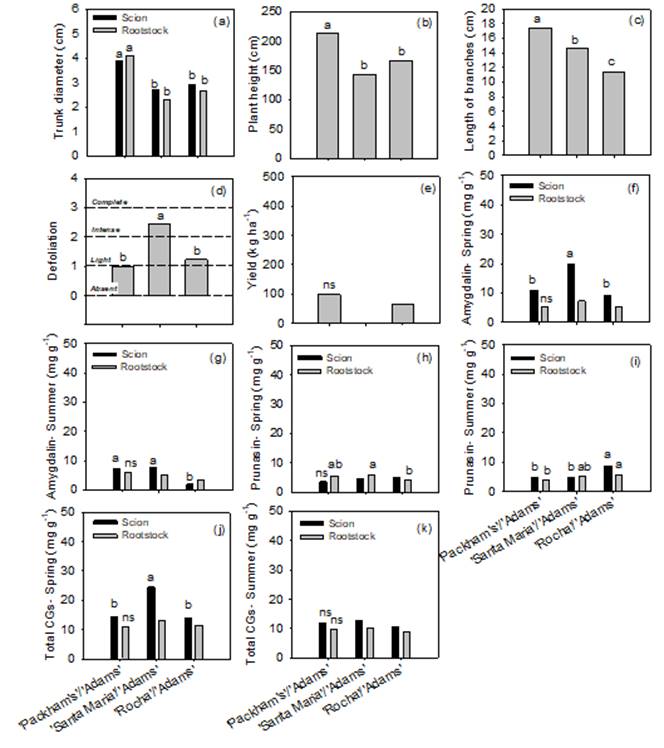
Figure 3. Growth, yield and biochemical assessments. *Different lowercase letters differ by Tukey’s test at 5 % probability of error.
The lower vegetative growth of Santa Maria/Adams combination is probably related to a lower affinity between graft components. The effect that is evident from vigor and graft compatibility assessment (Figure 4).

Figure 4. Vigour, graft incompatibility symptoms and biochemical assessments. *Vigor classes: ‘Very low’, ‘Low’, ‘Medium’, ‘High’ and ‘Very high’.Grafting incompatibility symptoms: ‘Absent’, ‘Light’, ‘Moderate’, and ‘Severe’. (ns) does not differ and (**) differs by Tukey’s test at 5 % probability of error.
The combination Packham’s Triumph/Adams was classified as ‘Medium’ vigor, while the others presented ‘Low’ vigor (Figure 4). As for the compatibility assessment, it was found that the combination Packham’s Triumph/Adams does not show any incompatibility symptoms, a result that is in line with what was observed by Pinto et al. (2009).
Rocha/Adams combination behaved similarly to Santa Maria/Adams for most growth variables but had less pronounced incompatibility symptoms, such as barely noticeable hypertrophy and little bud sprouting from the rootstock. Santa Maria/Adams showed clear symptoms of moderate incompatibility, with hypertrophy at the grafting point, in addition to a smaller trunk diameter on the rootstock and also a high emission of buds from the rootstock (Figure 4).
In the Santa Maria/Adams combination, the signs of grafting incompatibility are clear, but in relation to Rocha/Adams the evaluations carried out do not allow us to state that there is, in fact, grafting incompatibility. However, even if there is incompatibility of grafting in these two combinations, the degree must be light or moderate, with an impact only on growth control, inducing less vigor. Assumption supported by studies carried out in Vacaria-RS, in which ‘Rocha’ and ‘Santa Maria’ grafted on ‘Adams’ showed adequate growth and yield (Fioravanço et al., 2016; Fioravanço et al., 2017).
The three combinations studied presented low yield, even though significant flowering occurred (data not shown) (Figure 3e). In another region of Brazil, with higher altitude and cold accumulation, ‘Rocha’ and ‘Santa Maria’ grafted on ‘Adams’, showed a yield of 2.35 and 1.52 t ha-1, respectively, also in the third year after planting (Fioravanço et al., 2016; Fioravanço et al., 2017). Yield far superior to that observed in the present experiment. In addition to age, the low yield of the orchard evaluated in this study is likely to have been influenced by climatic factors, such as the lack of cold, as these cultivars need an average of 500-700 chilling hours (7.2 ºC) to break bud dormancy properly. However, the accumulation of chilling hours for this region, in that year, was only 270 hours, and in the last 3 years, the average was 362 hours. The insufficiency of cold in pear trees can result in greater floral abortion and, consequently, reduction in production (Pasa et al., 2012).
In the spring, ‘Santa Maria’ showed a higher amygdalin level than the other cultivars grafted on ‘Adams’ (Figure 3f). In summer, ‘Packham’s Triumph’ and ‘Santa Maria’ cultivars showed higher amygdalin levels (Figure 3g). Regardless of time year, there were no differences in the amygdalin concentration of rootstocks from studied combinations (Figures 3f and 3g). For prunasin in the spring, there were no differences between cultivars (Figure 3h). However, the rootstock ‘Adams’ had a higher concentration of prunasin when grafted under ‘Santa Maria’ (Figure 3h). In the summer, the combination Rocha/Adams showed a higher content of prunasin in the cultivar and rootstock, without differing only from the Rocha/Adams in the rootstock (Figure 3i).
In general, there was little difference between amygdalin and prunasin levels, except for the amygdalin in the ‘Santa Maria’, in spring (Figure 3f). This result was confirmed when evaluating the total CGs, when ‘Santa Maria’ presented a higher concentration in the spring is 2-fold higher than that determined in ‘Packham’s Triumph’ and ‘Rocha’ (Figure 3j). However, there were no differences between cultivars or rootstocks in the summer (Figure 3k).
In the analysis of the difference in concentration of total CGs between scion and rootstock of each combination, the main difference was between ‘Santa Maria’ and ‘Adams’, in which, ‘Santa Maria’ had a content of 11.29 mg g-1 or 1.9 times greater than ‘Adams’ (Figure 4). Difference that is probably related to the graft incompatibility symptoms and to the lower performance in the growth parameters of this combination. According to Pereira et al. (2018), a difference like this can cause severe graft incompatibility between Prunus persica and Prunus mume.
The biochemical results associated with the ‘Intense’ defoliation, the absence of fruit production, the ‘Low’ vigor and the incompatibility symptoms close to the grafting point, indicate that the incompatibility between ‘Santa Maria’ and ‘Adams’ exerts a moderate vigor control in this combination. A similar effect was reported by Pina et al. (2012) in Prunus combinations.
The Packham’s Triumph/Adams combination, on the other hand, proved to be completely compatible, with high level of vigor and similar CG levels between the graft components. However, for Rocha/Adams the occurrence of graft incompatibility is not clear, as some growth assessments indicate less vigor in relation to the Packham’s Triumph/Adams, but the symptoms of incompatibility considered in the present study do not allow a confirmation, as well a similar levels of CGs between scion and rootstock.
Cyanogenic glycosides x Grafting incompatibility between pear and quince
Among studied combinations, in both experiments, combinations with different levels of grafting incompatibility and CG concentrations were identified. Cascatense/P. Calleryana, Packham’s Triumph/Adams, and Rocha/Adams, had similar concentrations of CGs between scion and rootstock and showed no graft incompatibility symptoms. On the other hand, Santa Maria/Adams had ‘Moderate’ differences, resulting in less vigor and moderate incompatibility symptoms. But in the case of the Cascatense/EMC combination, the difference in the CG content between scion and rootstock was ‘High’ and the consequences were a ‘Severe’ incompatibility that compromised the growth and development of the combination.
In view of the results obtained in the present study, in which there is evident relationship between the content of CGs and the growth parameters, which in turn are related to the graft compatibility, a correlation study was carried out between the differences in the total content of CGs from scion and rootstock and the growth variables (Table 1). The results indicate that the difference in CG concentration between scion and rootstock in the spring really influences the growth of plants (trunk diameter, plant height, length of branches, and defoliation). This behavior is probably induced by the different levels of incompatibility of the studied combinations.

Conclusions
This study has shown that the graft incompatibility between pear and quince trees occurs in large part, due to the difference in the levels of CGs of both species. In addition, it can be suggested that when the difference between the contents of the scion and the rootstock is small (≤ 5 mg g-1), there is no effect on grafting compatibility; when the differences increase (> 5 and < 20mg g-1), an acceptable level of compatibility for the growth and development of the graft may occur but may result in the reduction of vigor at different levels. Finally, when the difference is high (≥ 20 mg g-1), in addition to the drastic reduction in vigor, there are problems of continuity in graft union, low yield, and early defoliation.
It is also added that the correct time for the assessment of CG levels is the beginning of spring, since it was at that moment that significant correlations between the levels of CGs of the evaluated genotypes and the growth of plants were identified (Table 1).
From the results presented in this study, a scheme that relates the difference in the concentration of CGs between scion and rootstock with the degree of graft compatibility (Figure 5) is proposed. This scheme will be based on the data obtained in the present study, in which five combinations will be evaluated, in two periods and in two experiments. Thus, it is believed that new studies should be performed, involving a greater number of combinations, in order to make this scheme more robust.


