











 English (pdf)
English (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mail Cited by SciELO
Cited by SciELO  Cited by Google
Cited by Google  Similars in
SciELO
Similars in
SciELO  Similars in Google
Similars in Google 
 Permalink
PermalinkIntroduction
Brazil is the second largest global producer of Coffea canephora, with the states of Espírito Santo and Rondônia as the main producers (Conab, 2020). This species has great economic and social importance in these regions, and scientific research on it covers various aspects such as genetic improvement focused on the development of cultivars, maintenance and expansion of genetic variability, and gaining an understanding of the reproductive system of this species (Ferrão et al., 2021).
Coffea canephora is an allogamous, diploid species with gametophytic self-incompatibility (Conagin and Mendes, 1961) that is readily propagated asexually (clonal) and sexually (seeds) (Berthaud, 1980). Since self-incompatibility, governed by the S gene, is determined by a prezygotic mechanism that influences the fertility rate, none of the allelic forms of the S gene are able to be fixed in a cross or population, thus resulting in heterozygous individuals only for this gene (Carvalho et al., 1983; De Nettancour, 1997; Silva et al., 2016; Moraes et al., 2018). Therefore, every plant is heterozygous for this gene because it is created from the union of two different gametes (Ferrão et al., 2017).
Pollination efficiency and fruiting are characteristics of the reproductive system to be considered in both the cultivation and breeding of this type of coffee. In these allogamous self-incompatible species, cross- fertilization occurs after the opening of flowers, and natural pollination is performed with the aid of wind and insects (Carvalho et al., 1983). As coffee has gregarious flowering, in natural conditions after the dry period, the first rains of the season cause all plants in a region bloom simultaneously with a variable number of flowers (Rena and Barros, 2004).
In C. canephora, the opening of flowers is completed in a few days. The flowers commonly open in the early hours of the morning, and their corolla begins to wither on the second day (Krug, 1937; Carvalho et al., 1983). Following this, the fruiting stage begins, which encompasses fruit set, development, and maturation.
According to Silveira and Carvalho (1996), the highest rate of fruit fall occurs at the end of the pinhead stage and during the rapid fruit expansion stage. Ronchi and DaMatta (2017) found that, beginning with an average of 24 pinheads per rosette, shortly after the initial stage of fruit expansion, this number was rapidly reduced to approximately 15 cherry fruits per rosette, resulting in a 38 % fruit drop during this period. The causes are related to pollination efficiency and environmental conditions, which influence on the plant’s ability to nourish new fruit.
Different methodologies have been explored for the transfer of desirable characteristics in this species using the existing genetic variability. The possibility of association of multiple favorable characteristics in a cultivar, with the manifestation of heterosis verified in crosses between contrasting parents, combined with the ease of use of this technique through clonal propagation, have established directed artificial hybridization as an important research tool. In the literature, it has been reported for some crops that the direction of the cross, whether direct or reciprocal, has an influence on the results (Ramalho et al., 2001).
Due to the self-incompatibility of the gametophytic type of the species, the compatibility of crossings of superior genetic materials and their consequences on the productivity and genetic variability of the offspring must always be studied (Ferrão et al., 2017). The fruiting percentage after directed hybridization varies according to the species and the environment, with average values of approximately 50 % (Carvalho et al., 1983; Rocha et al., 2015; Moraes et al., 2018; Souza et al., 2019). Using this approach, Charrier and Eskes (2004) reported that contamination is one of the most important sources of error in directed hybridization. Rocha et al. (2015), studying probabilities of errors in hybridization procedures, observed that the highest errors were related to the handling in the transfer of pollen grains between donor and receiving plants. According to the author, individuals with the rarest allelic forms of the S gene, which determines the expression of compatibility, benefit from the higher frequency of potentially compatible pollen-donor plants, making the fertility rate of the individual inversely proportional to the frequency of the allele in the population.
This work aimed to evaluate and compare the fruiting efficiency between reciprocal crosses in C. canephora using controlled hybridizations in the form of a full diallel with reciprocals.
Material and methods
Genetic materials
Controlled hybridizations in the field involving four parents, previously selected from the Incaper genetic improvement program, were performed at the Experimental Farm of Marilândia, located in Marilândia, ES, Brazil.
Genotypes (clones) 02, 03, 23 and 83 were selected for their favorable and complementary agronomic characteristics, considering the hybridization between plants of the Conilon and Robusta groups. Clones 02, 03, and 23 had characteristics typical of the Conilon group and clone 83 had characteristics typical of the Robusta group. The genetic materials of the Conilon group differ from those of Robusta by having plants with lower growth, a greater number of smaller-diameter branched stems, smaller elongated leaves, an earlier maturation cycle, greater tolerance to drought, and greater susceptibility to the most common coffee diseases (Ferrão et al., 2019). Regarding the maturation time, clone 23 was the earliest, followed by clones 02, 03, and 83, the last of which had medium maturation.
Hybridization was performed in a full diallel design with reciprocals, with four replicates. In this design, the number of crosses is determined by the expression NC = N × (N-1) × r, where NC is the number of crosses, N is the number of parents (4), and r is the number of repetitions (4), resulting in a total of 48 directed hybridization procedures (TNC = (4 × (4-1)) × 4 = 48).
Thus, 12 treatments (crossings) were evaluated: 02 x 03, 03 x 02, 02 x 23, 23 x 02, 02 x 83, 83 x 02, 03 x 23, 23 x 03, 03 x 83, 83 x 03, 23 x 83, and 83 x 23.
Directed hybridizations
The hybridizations were carried out in the field, within the Active Germplasm Bank Experiment, with the genotypes cultivated in plots of 10 plants. In each plot of the corresponding treatments, four plants were marked as female parents and four plants as male parents.
In each repetition of the cross, one branch per plant was used, with the protection of the entire plagiotropic branch using the total of rosettes with unopened flowers. The day before anthesis, plagiotropic branches with white flower buds were protected using 55 × 25 cm kraft bags. Pollinations were carried out from 07:00 to 09:30, a period of high pollen grain abundance. Donor and receiving branches were selected from among those with inflorescences in the same phenological stage. Crosses were performed from August 28 to August 30.
An opening was made at the tip of the paper bag to contact the donor and receiver branches. After the contact between the inflorescences, the procedure was ended by closing the paper bag and identifying the cross.
Evaluations of fruit development
The dormancy stage lasted 90 days and began after artificial hybridization, when it was not possible to observe fruit growth. After this period, fruit development and growth were evaluated. Abscission of unfertilized fruits occurred soon after the dormancy period, and a low abscission of fruits was observed throughout the maturation period. Fruit growth was monitored by visual evaluation at the pinhead, dried, green, and cherry stages.
The following characteristics were evaluated: TNP: total number of pinheads (initial fruit set) per cross evaluated 26 days after crossing, NGF: number of green fruits per cross evaluated 125 d after crossing, NCF: number of ripe cherry fruits per cross evaluated at the time of ripe fruit harvest, FS: fruit set, evaluated as a percentage using the equation (NGF/ TNP) × 100, and FS1: fruit set between the number of ripe cherry fruits and green fruits, evaluated in percentage by the equation (NCF/NGF) × 100.
Statistical analysis
The characteristics were interpreted considering the analysis of individual variance, means comparison test, estimates of genetic parameters and correlations, and the effect of genotypes as fixed. Statistical analyses were performed using GENES 2016 software (Cruz, 2016).
Results
In the analysis of variance, there was a significant effect of the source of variation of treatments on all characteristics, according to the F test at the 1 % or 5 % probability level (Table 1). The estimates of the environmental coefficient of variation (CVe) were high for the number of green and cherry fruits per branch, and the ratio between the genetic and environmental coefficients of variation (CVg/CVe) was greater than 1.0 for the total number of pinheads per branch and the percentage of fruit set (fruiting) only. These characteristics had estimates of genotypic coefficient of determination (H) higher than 80 %, characterizing good experimental precision and favorable conditions to obtain gains with selection.
Table 1 Summary of analysis of variance, estimates of genetic parameters and comparison of means for six traits, obtained from 12 hybrid combinations of
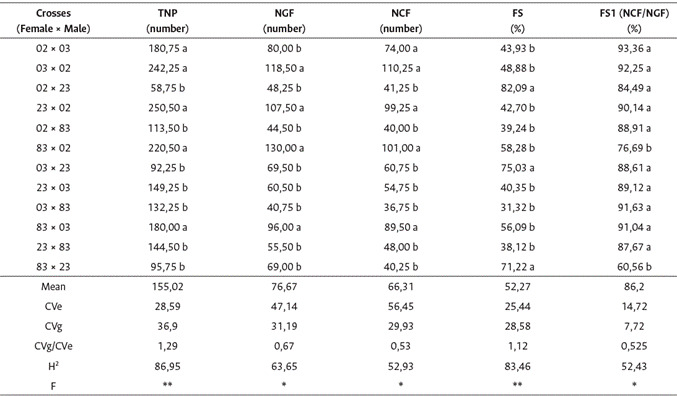
Means followed by at least one letter in the column did not differ from each other by the Scott-Knott test (p ≤ 0,05). CVe = experimental coefficient of variation; CVg = genetic coefficient of variation; H2 = genotypic coefficient of determination; F = F test of the analysis of variance associated with hypothesis of existence of genetic differences between treatments; TNP = total number of pinheads per cross; NGF = number of green fruits per cross; NCF = number of ripe cherry fruits per cross; FS = fruit set; FS1 (NCF/NGF) = ratio between the number of ripe cherry fruits and green fruits.
In the comparison of means, important differences were observed between treatments in the direct and reciprocal hybridizations. When clone 23 was used as the male parent, 58,75 % to 95,75 % of the pinheads (TNP) were observed, as well as the highest fruiting rates (FS), ranging from 71,22 % to 82,09 % (Table 1). In contrast, clone 83 was the male parent with the lowest fruiting rate (31,32 % to 39,24 %).
Crosses using clone 02 as the male parent showed the highest number of pinheads, green fruits, and ripe cherry fruits per branch, and the average fruiting rate ranged from 42,70 % to 58,28 % in the three combinations. The highest ratios between ripe cherry fruits (NCF) and green fruits (NGF) involved clone 03, and the lowest ratios were observed in crosses 83 × 23 and 83 × 02. On average, this ratio was 86,2 % for the FS1 (NCF/NGF).
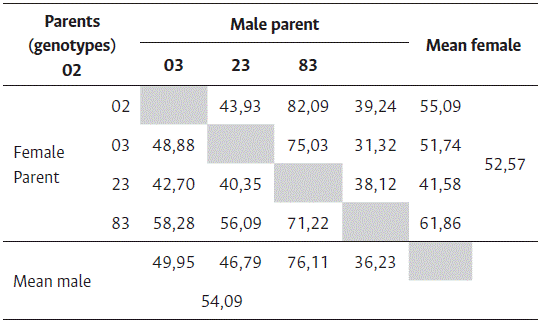
The joint data of fruit set showed an average of 52,27 % with an amplitude from 31,32 % to 82,09 %, and significant differences between some reciprocal crosses (Figure 1). The fruiting rate of male parents was 54,09 % and that of females was 52,57 %, showing the average non-superiority of genotypes when used as parents (Table 2).
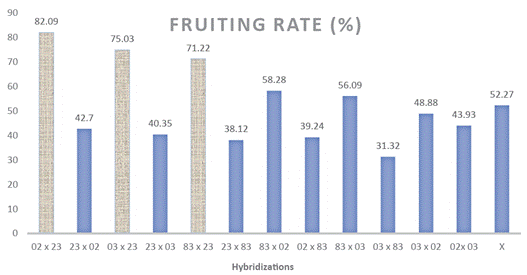
Figure 1 Fruit set of 12 hybrid combinations and the overall mean (x) estimated using a full diallel design with reciprocals among four parents with characteristics of the Conilon and Robusta groups, at the Experimental Farm of Marilândia, ES, Brazil. In 02 x 23 crosses, the first is the female parent (02) and the second is the male parent (23).Means followed by the same color (columns) did not differ from each other by the Scott-Knott test (p ≤ 0,05).
Table 2 Means of fruit set (%) using the four genotyps of Coffea canephora as male and female parents, in a diallel trial with the reciprocals, at the Experimental Farm of Marilândia, ES, Brazil
It was verified that the estimates of genotypic correlations generally surpassed those of phenotypic correlations and were negative for all associations involving the fruiting rate characteristic (Table 3), being of greater magnitude with the total number of pinheads (-0,54). The characteristic total number of pinheads showed positive, high genotypic and phenotypic correlations with the number of green and ripe cherry fruits. As expected, the highest correlations were between the number of green fruits and the number of ripe cherry fruits, with values above 95 %. For some characteristics, the environmental correlations showed differences in magnitude and signal, such as for NGF, FS, NCF, and FR, indicating that direct selection, based on genetic correlation coefficients, may be impaired by the differential action of the environment on the variables involved, which may favor one and disfavor the other.
Table 3 Estimates of phenotypic, genotypic, and environmental correlation coefficients corresponding to the combinations of five characteristics evaluated in 12 hybrid combinations of Coffea canephora, at the Experimental Farm of Marilândia, ES, Brazil
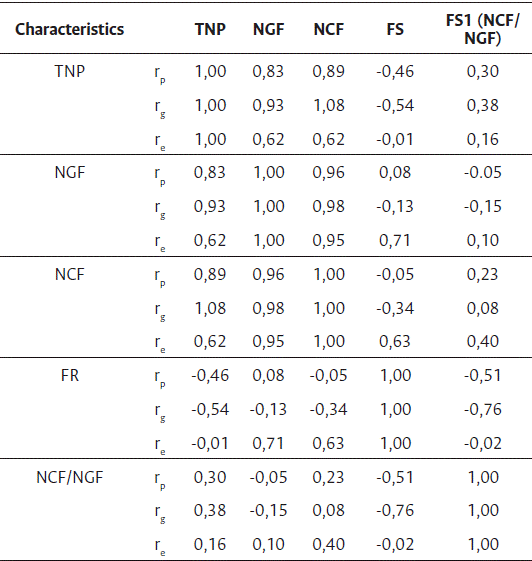
*rp = phenotypic correlation coefficient, rg = genotypic correlation coefficient, re = environmental correlation coefficient. TNP = total number of pinheads/branch; NGF = number of green fruits/branch; NCF = number of ripe cherry fruits/branch; FS = fruit set (%) and FS1 =NCF/NGF = ratio between number of ripe cherry fruits and green fruits (%).
Discussion
The estimates of the genetic parameters indicated a predominance of the genetic component in the expression of total number of pinheads per branch (TNP) and percentage of fruit (FS), indicating that the effect of the genotypes outweighed the effect of the environment on the expression of this characteristic.
In this study, differences were observed between reciprocal crosses involving genotypes 83 and 23, highlighting the maternal effect for the combinations in which these parents are used as pollen-receiving plants.
According to Ramalho et al. (2001), most of the characteristics of higher organisms are controlled by nuclear genes, which segregate according to the behavior of chromosomes during meiosis. However, there is another group of characters inherited as a function of genes or gene products present in the cytoplasm, in which the female gamete contributes almost entirely to the descendant. This type of inheritance can be explained by the maternal effect or extrachromosomal inheritance, and for their identification it is necessary to verify whether there is a difference between the results of a cross and its reciprocal. Ramalho et al. (2001) state that if nuclear genes control the inheritance of a particular character, the results of a cross and its reciprocal will be similar. If there are cytoplasmic effects, the results of reciprocal crosses will be different, and the phenotype of the descendants will be influenced by the female parent, which contributes to the cytoplasm.
Based on the average behavior of the clones for FS, crosses involving clones 02 and 03 showed no statistical differences when used reciprocally, and their average FS was 50,89 %. In turn, clone 23 was the donor plant with the superior fruiting rate (76,11 %) and clone 83 emerged as the best receiving plant (61,86 %). In the comparison of these two genotypes (23 and 83), antagonism was observed in their behavior when used as pollen-donor plants (male parents) and pollen-receiver plants (female parents). This contrast between direct and reciprocal crosses is defined as a maternal effect and has great importance for species improvement, which is based on the understanding of the reproductive system of coffee as a method of utilizing and manipulating genetic variability. Hybridization between Conilon and Robusta parents with contrasting and complementary characteristics seeks the production of new hybrid plants that prevail for the expression of the best characteristics of the botanical groups associated with the vigor of the hybrid (Oliveira et al., 2018).
Moraes et al. (2018) described that individuals with the rarest allelic forms of the S (incompatibility) gene benefit from a higher frequency of potentially compatible pollen-donor plants, making the individual’s fertility rate inversely proportional to the frequency of the allele in the population. Thus, it is inferred that among the four recombined genotypes in this study, clone 23 had the most divergent allelic forms. Additionally, it is worth mentioning that the fruiting rate found in the present study is in accordance with other studies, such as Moraes et al. (2018), who found that the compatible crosses had a mean rate of 44 %, with a range from 26 % to 77 %. The same authors found that in the incompatible crosses, the rate of 3 % was caused by contamination during the hybridization procedure.
Different studies on the line of self-incompatibility with Coffea canephora confirm that it is gametophytic, governed by the action of a single S gene and comprised of three to four allelic forms (S1, S2, S3, S4) (Conagin and Mendes, 1961; Berthaud, 1980; Lashermes et al., 1996; Moraes et al., 2018). Thus, three levels of compatibility between plants are expected: self-incompatible pollination, when the parents have identical allelic forms (S1S2 × S1S2); partial compatibility, where the parents have a common allelic form (S1S2 × S1S3); and total compatibility, when the parents have different allelic forms (S1S2 × S3S4) (Ferrão et al., 2017). However, there are many questions regarding partially compatible crosses. According to Moraes et al. (2018), it can be mistakenly assumed that the result of partially compatible crosses is 50 % fertilized locules and 50 % unfertilized locules. However, these proportions do not occur because self-incompatibility is a prezygotic mechanism, and pollination of non-compatible pollen grains does not prevent the germination and development of pollen tubes of compatible grains. Therefore, directed crosses between two partially compatible plants can naturally result in fertilization rates higher than 50 %, and the percentage of occurrence of peaberry grains cannot be used to infer the genotype of the S gene, since the efficiency of pollination depends on the transport, germination, and development of the pollen tube of compatible pollen grains.
In the study of the correlations, it was found that the estimates of the genotypic correlations (rg) were negative for all associations involving the characteristic fruit set (FS), being of greater magnitude for the association with total number of pinheads/branches (TNP), indicating that a larger number of pinheads negatively interferes with the fruiting rate. The greater magnitudes of the estimates of the genotypic correlations found compared to the phenotypic correlations, for most of the analyzed combinations, showed a stronger influence of the genetic components on the expression of characteristics associated with the development and growth of the fruits.
The fruit maturation is influenced by both genetic and environmental factors that impact the processes of flowering and fruit development. During the flowering phase, irregularity in rainfall distribution can lead to longer intervals between the main blooms. This was observed by Souza et al. in 2017, who identified a greater disparity in fruit maturation caused by an interval of more than 40 days between the two main flowerings in the Amazon region. In addition to environmental factors, genetic traits also play a significant role. There are genotypes that exhibit distinct responses during the flowering process. For instance, the BRS2299 clone stands out due to its high rate of flowerings throughout the year, as observed by Moraes et al. in 2018. Overall, greater maturation uniformity is observed when rainfall is well-distributed during the flowering period. Our results indicate that only hybridizations using genotype 83 as the female receptor (83 x 23 and 83 x 02) resulted in lower fruit maturation uniformity (Table 1).
Conclusions
The estimates of the genotypic coefficients of determination at the mean level (H2) and the ratio between the genetic and environmental coefficients of variation (CVg/CVe) were higher than 83,46 % and 1.0, respectively, for the total number of pinheads per cross (TNP) and fruiting rate (FS), indicating good experimental precision and genetic variability for the selection.
The mean fruit set was 52,27 %, ranging from 31,32 % to 82,09 %. There were differences between the reciprocal crosses, evidencing a possible maternal effect for some combinations, which should be investigated in further work.
Clone 23 was the strongest male parent and weakest female parent, while clone 83 emerged as the superior female parent and inferior male parent. In crosses involving clones 02 and 03, there were no significant differences when used reciprocally, and the average FS was 50,89 %. Clone 23 predominated as a pollen donor, which indicates the occurrence of rarer allelic forms of the S (incompatibility) gene among the evaluated parents.
The estimates of genotypic correlations were negative for all associations involving the FS characteristic, being of greater magnitude with TNP (-0.54).

