











 Spanish (pdf)
Spanish (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mail Cited by SciELO
Cited by SciELO  Cited by Google
Cited by Google  Similars in
SciELO
Similars in
SciELO  Similars in Google
Similars in Google 
 Permalink
PermalinkINTRODUCCIÓN
En la actualidad, México cuenta con más de 100 sitios reconocidos como humedales de importancia internacional por la Convención de Ramsar. La Ciénaga de Tamasopo o Ciénega de Tampasquín en San Luis Potosí es un humedal léntico con clima neotropical. Sus características ecológicas le permiten albergar especies de flora y fauna endémicas desde plantas hiperacumuladoras hasta una variedad de anfibios, reptiles, aves e invertebrados (Torres, 2008); el humedal fue declarado por la Convención Ramsar en 2008 como de importancia internacional. En los últimos años, los suelos de este ecosistema se han visto modificados en tierras de uso agrícola, principalmente, para siembra de caña de azúcar (Torres, 2008; Tapia-Goné etal., 2010; Pérez-Castillo et al., 2017. Fig. 1a), donde comúnmente se utilizan herbicidas e insecticidas como glifosato, atrazina, paraquat o clorpirifos, metanoarsonato monosodico, entre otros (Díaz y Labrada, 2016). De acuerdo con información proporcionada (comunicación personal) por la Unión Local de Productores de Caña de Azúcar (CNC) de Ciudad Valles y de Tamasopo, la aplicación de plaguicidas en cultivos de caña de azúcar en la Huasteca Potosina (Fig. 1b) se da a lo largo de todo el año, existiendo dos temporadas de aplicación: una alta, que corresponde a los meses de junio a septiembre y una baja, que corresponde a los meses de noviembre a febrero.

Figura 1 Cultivos de caña de azúcar próximos al humedal (a), aplicación de plaguicidas en los cultivos (b), disposición de recipiente de plaguicidas (-glifosato, ácido 2,4-diclorofenoxiacético-) próximos a los cuerpos de agua (c), individuos de caracoles de agua dulce (d).
Una de las prácticas que más concierne con esta problemática es la utilización de biocidas para el control de plagas y malezas indeseables en los cultivos, debido a que, ya sea directa o indirectamente, pueden afectar a los organismos no objetivo de los ecosistemas aledaños (Albert y Benítez, 2005; Albert y Loera-Gallardo, 2005; Sánchez-Bayo et al., 2011. Fig. 1c). En este ámbito, en algunas ocasiones es difícil evaluar las concentraciones de plaguicidas debido a que algunos pueden degradarse rápidamente, por lo que sus residuos pueden estar bajo el límite de detección de los aparatos empleados para cuantificarlos (Chitmanat et al., 2008). Ante esto, la evaluación de alteraciones enzimáticas, proteicas, de energía u hormonales (biomarcadores) se ha convertido en una herramienta útil para evidenciar y monitorear la exposición y efectos subletales de plaguicidas en organismos, debido a que funcionan como una señal de alerta temprana para prevenir consecuencias a futuro o en niveles mayores de organización biológica (Depledge, 1994; Amiard-Triquet et al., 2013).
En algunos estudios, en escenarios reales y de laboratorio, se han utilizado biomarcadores expuestos a plaguicidas; este es el caso de la inhibición de acetilcolinesterasa (AChE), el aumento de glutatión s-transferasa (GST) y la presencia de malondialdehído (MDA) en organismos como los macroinvertebrados (ej. gasterópodos) (Singh et al., 1993; Chitmanat et al., 2008; Domingues et al., 2010; Rivadeneira et al., 2013; Basopo y Naik, 2015). La AChE es una enzima responsable de la señal de sinapsis colinérgicas mediante la hidrólisis del neurotransmisor acetilcolina. El resultado de una exposición a plaguicidas organofosforados y carbamatos conlleva por lo general a una inhibición de esta enzima y a efectos que pueden resultar adversos para los organismos (Thompson, 1999). Por su parte, la GST es una enzima perteneciente a los procesos de biotransformación fase II para eliminar xenobióticos de un organismo; se encarga de combinar glutatión reducido con compuestos electrófilos (xenobióticos conjugados) formando tioéteres que pueden ser más fácil de eliminar del organismo. El aumento de esta enzima en tejido o plasma de un organismo es comúnmente indicativo de una exposición a compuestos hidrocarburos, plaguicidas y bifenilos policlorados (Slatinská et al., 2008). Por otra parte, el MDA es un aldehido producto final e indicativo de la peroxidación lipídica, esta última considerada como uno de los mecanismos que ocasionan daño oxidativo a membranas celulares y procesos de toxicidad que conducen a la muerte celular (Repetto et al., 2012). Por último, se ha demostrado que además de la exposición a plaguicidas, existen diversos factores extrínsecos (ej. parámetros fisicoquímicos del agua, temporalidad, etc.) que influyen en la variación de estos biomarcadores en organismos como los moluscos (Amiard-Triquet y Brigitte Berthet, 2014).
Los moluscos tienen un papel muy importante en los ecosistemas de humedal; son componentes clave en la cadena trófica, debido a que vinculan la materia orgánica y los recursos nutritivos con los niveles más altos de la cadena; además, se consideran como bioindicadores de la salud de este tipo de ecosistemas (Oehlmann y Schulte-Oehlmann, 2003; Li et al., 2010). Los gasterópodos son un grupo de moluscos que se han utilizado como bioindicadores para evaluar la exposición a metales pesados, plaguicidas y bifenilos policlorados (Chitmanat et al., 2008; Domingues et al., 2009; Radwan et al., 2010; Fu et al., 2011; Grara et al., 2012; Cossi et al., 2015). En México, se destaca la familia Pachychilidae de caracoles de agua dulce (Pachychilus sp.), los cuales se han considerado bioindicadores de la contaminación de ecosistemas acuáticos (Naranjo-García y Meza Meneses, 2011. Fig. 1d). La distribución geográfica nativa del género Pachychilus se restringe a países como Guatemala, Brasil, Nicaragua, Belice, Cuba y México (Pérez y López de la Fuente, 1993; Vázquez-Perera y Perera-Valderrama, 2010; Gómez-Berning et al., 2012).
En este contexto, el objetivo de esta investigación fue evaluar la actividad de biomarcadores de exposición a plaguicidas (AChE, GST y MDA) en dos temporadas (secas y post-lluvias) en tejido de gasterópodos (caracol de agua dulce -Pachychilus sp.-) residentes de sitios impactados por la agricultura en la Ciénaga de Tamasopo (S.L.P., México).
MATERIALES Y MÉTODOS
Sitio de estudio y colecta de organismos
La Ciénaga de Tamasopo se ubica en el estado de San Luis Potosí; México (Fig. 2) en las coordenadas 99°19'14,00'' E, 99°16'57,00'' W, 21°52'03,00" N y 21°45'41,00'' S. El muestreo se llevó a cabo en dos temporadas: marzo (secas) y noviembre (post-lluvias) del 2016. Se eligieron tres estaciones de muestreo a lo largo del humedal de acuerdo con estudios previos en la zona (Pérez-Castillo et al., 2017). Cabe resaltar que estas estaciones fueron utilizadas en cada temporada de muestreo. La primera estación de muestreo (Mirador -MIR-) se localizó en las coordenadas 21°49'0,47'' N y 99°18'15,58'' W, la segunda estación (Capuchinas -CAP-) se ubicó en las coordenadas 21°46'15,14'' N y 99°17'46,03'' W, y la tercera estación (Ejido Veinte -VEI-) tuvo las coordenadas 21°45'15,14'' N y 99°16'48,77'' W. La estación MIR se consideró como la de menor influencia agrícola en comparación con las estaciones CAP y VEI, las cuales tuvieron mayor influencia de cultivos aledaños de caña de azúcar.

Para la colecta de los caracoles de agua dulce, se realizaron trayectos lineales de 200 m recolectando especímenes de Pachychilus sp.; posteriormente se seleccionaron de forma aleatoria diez organismos por estación de muestreo, dando un total de 30 organismos por temporada. Se seleccionaron organismos adultos con una talla de 32,6 ± 2,14 mm (de 27,9 a 37,4 mm) y un peso de 1,67 ± 0,58 g (de 0,6 a 2,8 g), así como de cinco a seis vueltas de la concha y ápices desgastados. Los organismos no se seleccionaron por sexo debido a que existe poca información sobre las características externas dimórficas para la especie (Naranjo-García y Meza-Meneses, 2011). Después de su colecta, los caracoles se sacrificaron y se les extrajo el tejido total de la concha. Debido al pequeño tamaño de los ejemplares, se optó por utilizar todo el tejido del organismo. Los tejidos se conservaron en nitrógeno líquido (-186 °C) para su transporte final al laboratorio. La colecta se realizó bajo el permiso FAUT-0321 otorgado por la Secretaria de Medio Ambiente y Recursos Naturales (SEMARNAT). La identificación taxonómica se realizó a nivel de género de acuerdo con información documental (Burch y Cruz-Reyes, 1987; Wong-Argüelles, 2005; Naranjo-García y Meza-Meneses, 2011). La identificación a nivel de especie no se pudo realizar debido a que los caracoles retienen características morfológicas primitivas (Healy et al., 1990); además, hay un gran número de endemismos en la zona que aún no están descritos.
Adicionalmente, en cada sitio de colecta se determinaron la temperatura, pH, oxígeno disuelto y conductividad eléctrica del agua con una sonda multiparamétrica (HANNA® instruments, HI 9828). Las mediciones se realizaron entre 1 y 1,5 m de profundidad en los trayectos de colecta de organismos, con lapsos de medición no mayor de una hora entre estaciones de muestreo.
Determinación de biomarcadores
En el laboratorio, la homogenización de los tejidos totales de caracol se realizó con buffer tris-acetato 10 mM pH 7,4, que contenía aprotinina (como inhibidor de proteasas), 140 mM de NaCl y 1 mM EDTA, a 1000 rpm a 4 °C con un homogenizador Polytron (OMNI International GLH), en una proporción 1:4 (tejido: buffer). Posteriormente, se obtuvo la fracción s12 (sobrenadante) de los homogenados por centrifugación a 12 000 g x 20 min a 4 °C.
La determinación de acetilcolinesterasa se basó en el método de Ellman et al. (1961) modificada por Gagné (2014) para microplaca de 96 pozos, donde 50 µL de sobrenadante (s12, previamente diluido) se mezclaron con 100 µL de acetiltiocolina (1 mM) y 100 µL de reactivo de Ellman (1 mM) en una microplaca a 30 °C, y se midió la cinética por 10 min a 412 nM en un lector de microplacas UV-Visible (BioTek® Sinergy H1). Para el blanco analítico se utilizó buffer tris-acetato. Lo mismo se realizó con un estándar de tiol con una concentración de 25 mM en un volumen de 250 mL. Para el cálculo de la concentración de AChE se utilizó la siguiente fórmula:

Donde ∆Abs= cambio de la absorbancia (10 min - 0 min), conc. estándar= 25 nM/mL de tiol, FD= factor de dilución (5).
Para la cuantificación de la GST se utilizó el método propuesto por Habig et al. (1974) modificado por Gagné (2014) para microplaca de 96 pozos; se mezclaron 50 µL de sobrenadante (s12, previamente diluido) con 100 µL de glutatión reducido (GSH, 1 mM) y 100 µL de sustrato 1-cloro-2,4-dinitrobenceno (CDNB, 1 mM) en una microplaca y se midió la cinética por 10 min a 340 nM en un lector de microplacas UV-Visible (BioTek® Sinergy H1). Como blanco analítico se utilizó buffer fosfato. Para el cálculo del rango de conjugación de GST se utilizó la siguiente fórmula:

Donde ∆Abs= cambio de la absorbancia (10 min - 0 min), FD= factor de dilución (5).
La determinación y el cálculo de malondialdehído en el sobrenadante s12 de caracoles se realizó siguiendo las instrucciones del kit comercial "NWLSS™ Malondialdehyde Assay Single Point" (Cat. No. NWK-MDA01, NWLSS, EUA) para espectrofotometría UV-Visible. El sobrenadante s12 se mezcló con soluciones de ácido tiobarbitúrico (TBA), hidroxitolueno butilado (BHT) y ácido fosfórico, y se incubó a 60 ° C para formar el aducto MDA-TBA2 que absorbe fuertemente a 532 nm. La concentración de MDA para cada muestra se obtuvo a partir de la interpolación de la absorbancia en una curva de calibración hecha con un estándar de tetrametoxipropano (1-4 mM) propia del kit comercial.
Las concentraciones de los biomarcadores se determinaron por triplicado y se normalizaron por la cantidad de tejido húmedo utilizado (g de tejido/mL de buffer de homogenización). Esto último se realizó dividiendo la actividad de las enzimas o la presencia de MDA por el tejido fresco utilizado para la determinación conforme a lo descrito por Gagné (2014).
Todos los reactivos fueron de grado bioquímico y adquiridos de las casas comerciales SIGMA y Northwest Life Science Specialities™.
Análisis estadístico
Se realizó una prueba de bondad de ajuste de Kolmogorov-Smirnov para determinar la normalidad de los datos. Dado que los datos no siguieron una distribución normal, se realizaron pruebas no-paramétricas. Para las comparaciones entre los biomarcadores por temporadas (seca y post-lluvias) se utilizó la prueba de U de Mann-Whitney (U-MW), mientras que las comparaciones entre estaciones de muestreo por temporada se hicieron mediante un análisis de Kruskall-Wallis (KW-H). Además, se hicieron asociaciones entre los biomarcadores por temporada, las cuales se evaluaron con una correlación de Spearman una vez que los datos fueron transformados logarítmicamente. La significancia estadística se evaluó en 1 % y 5 % con el software Prisma GraphPad 6.0. Los datos se reportaron como la mediana y el rango intercuartil, y se complementaron con los mínimos y máximos.
RESULTADOS
Los parámetros fisicoquímicos determinados en agua de la Ciénaga de Tamasopo fueron similares para ambas temporadas, a excepción de la conductividad eléctrica del agua, que se encontró mayor (51 %) en la temporada de secas con respecto a la temporada de lluvias (Tabla 1).
Tabla 1 Parámetros fisicoquímicos medidos por temporada en la Ciénaga de Tamasopo, S.L.P., México.

media ± error estándar
En cuanto a los biomarcadores no se encontraron diferencias entre las estaciones de muestreo a lo largo del humedal, pero si entre temporadas. Al respecto, la actividad de AChE en el tejido de los caracoles varió de 23,6 a 210,5 nM x min-1 x g de tejido; en temporada de secas fue mayor (39,0 %) con respecto a la que se obtuvo en post-lluvias (U-MW30 30 = 2 5 2, p<0,05, Fig. 3a). A su vez, la actividad de GST varió de 74,1 a 303,5 abs x min-1 x g de tejido, siendo un 28,5 % mayor en la temporada de secas con respecto a la temporada de post-lluvias (U-MW3030= 1 1 3, p<0,001, Fig. 3b). Los valores de MDA fueron de 3,2 a 41,6 mM x g de tejido; se observó una mayor presencia (25,0 %) de este biomarcador en la temporada de post-lluvias (U-MW30 30= 252, p<0,05, Fig. 3c).
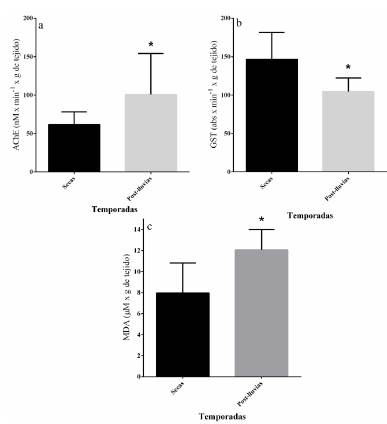
Figura 3 Actividad de AChE (a) y GST (b), y presencia de MDA (c) en tejido de caracoles de agua dulce (Pachychilus sp.) de la Ciénega de Tamasopo por temporadas, prueba de U-Mann-Whitney, *p< 0,05. Se presentan la mediana y rango intercuartil. Secas (n = 30) y Post-lluvias (n = 30).
Por último, se encontraron asociaciones entre biomarcadores en ambas temporadas (Tabla 2). En la temporada de secas se observó una correlación estadísticamente significativa (r = 0,42, p< 0,05, n = 30) entre la actividad de AChE y la de GST (Fig. 4a). En la temporada de post-lluvias se obtuvo una correlación estadísticamente significativa (r = 0,60, p< 0,001, n = 30) entre la concentración de MDA y la actividad de GST (Fig. 4b).


DISCUSIÓN
Los caracoles de agua dulce (Pachychilus sp.) de la Ciénaga de Tamasopo presentaron diferencias entre temporadas en los biomarcadores evaluados, lo cual podría deberse a diferentes factores. El primero de estos puede estar relacionado con los parámetros fisicoquímicos en el agua y el segundo con una exposición a plaguicidas utilizados en los cultivos alrededor del humedal. En cuanto al primer factor, considerando los criterios ecológicos de calidad de agua (CE-CCA-001/89, SEDUE, 1989) para la prevención de la contaminación del agua y la protección de la flora y la fauna acuática de los cuerpos de agua dulce en México, los parámetros fisicoquímicos del agua se encontraron dentro de los valores establecidos como normales; además, la mayoría de estos registros fueron similares en ambas temporadas de muestreo. Los valores de los parámetros en el agua son muy similares a los obtenidos por Pérez-Castillo (2018) en este mismo sitio de estudio, así como, en los de otros humedales intercontinentales en el mundo (Gerla, 2013; Barman et al., 2015). Sobre los efectos de otros parámetros como el oxígeno disuelto y el pH sobre los biomarcadores, la información es muy limitada y únicamente se ha relacionado en grupos como bivalvos y crustáceos (Amiard-Triquet y Berthet, 2014).
Es importante resaltar que la conductividad eléctrica fue mayor en la temporada seca; este parámetro está relacionado directamente con la salinidad, debido a que una cantidad mayor de iones disueltos en el agua aumenta su capacidad de conducir electricidad. Se ha demostrado que los cambios de temperatura y salinidad pueden influir en la actividad de biomarcadores como AChE, GST y de estrés oxidativo, lo cual se ha documentado mayormente en moluscos bivalvos (Pfeifer et al., 2005; Fossi-Tankoua et al., 2011; Gélinas et al., 2013). De manera general, se ha registrado un decremento de la actividad de AChE y un aumento en la actividad de GST en bivalvos, cuando la salinidad aumenta (Damiens et al., 2004; Pfeifer et al., 2005; Fossi-Tankoua et al., 2011), siendo este patrón similar al obtenido en la presente investigación; otro tanto puede decirse de la asociación encontrada entre estos dos biomarcadores. Por lo anterior, es posible que las alteraciones de los biomarcadores encontrada entre temporadas en Pachychilus sp. de la Ciénaga de Tamasopo puedan también ser atribuibles a los cambios de salinidad en el agua del humedal. La disminución de AChE podría estar directamente relacionada con el cambio en la concentración de iones inorgánicos (extra e intracelulares) que ocurre durante la adaptación a las variaciones osmóticas en los organismos osmoconformadores y que afectan con la capacidad de hidrólisis del sustrato (Tomlinson et al., 1981; Pfeifer et al., 2005). Adicionalmente, el incremento de GST podría estar mediado por condiciones moderadas de salinidad y podría tener un efecto protector en el organismo ante el estrés oxidativo al mejorar las condiciones fisiológicas de producción enzimática (Zanette et al., 2011). Por otro lado, también se ha relacionado un incremento en la solubilidad y la biodisponibilidad de los contaminantes orgánicos en fusión de la salinidad del agua (Zanette et al., 2011).
El segundo factor al que se le puede atribuir las diferencias en los biomarcadores entre temporadas, es la exposición a plaguicidas. Estos pueden ocasionar respuestas en los organismos a los cuales no van dirigidos (Reylea y Hoverman, 2008; Sánchez-Bayo et al., 2011). El hecho de encontrar una menor actividad de AChE y un mayor rango de conjugación de GST en temporada seca, inverso a lo ocurrido en la temporada de post-lluvias, puede atribuirse al patrón de aplicación de plaguicidas en los cultivos aledaños al humedal y al proceso de concentración de contaminantes en temporada seca. La GST es una de las enzimas que conforman la segunda fase de biotransformación de xenobióticos, conjugando estos compuestos para que puedan ser eliminados del organismo (Parkinson et al., 2013). El aumento del rango de conjugación de GST y la menor actividad de AChE en la temporada seca parecen asociarse a la exposición a los plaguicidas aplicados en el humedal; adicionalmente, se encontró una asociación entre las dos enzimas lo que evidencia la compensación de los organismos ante este estrés. Al respecto, diferentes estudios en organismos invertebrados han observado el mismo patrón encontrado en este estudio (Moreira y Guilhermino, 2005; Domingues et al., 2009; Vidal-Liñán et al., 2014).
Se ha demostrado que la GST también puede cumplir un papel como antioxidante y como mecanismo de defensa ante la peroxidación de lípidos. En general, la GST puede reducir los radicales peróxidos formados de ácidos grasos insaturados, así como los productos finales (4-HNE, MDA), e interrumpir la cadena que se origina por la peroxidación lipídica (Yang et al., 2003; Sharma et al., 2004). En nuestro caso, la dinámica hídrica en temporada de post-lluvias (ej. resuspensión de sedimentos) podría incrementar la cantidad de contaminantes con potencial de generación de radicales libres en el sistema acuático (compuestos orgánicos persistentes, metales pesados, hidrocarburos aromáticos policíclicos, etc.), lo que incrementaría el daño en los organismos.
Algunos estudios en bivalvos han demostrado una mayor peroxidación lipídica y una disminución en los antioxidantes o viceversa, como mecanismos de compensación ante un estrés oxidativo, lo cual es congruente con los resultados obtenidos en este estudio, sobre todo con la correlación encontrada entre MDA y GST (Fig. 4b). Muy pocos estudios se han hecho con la peroxidación de lípidos como biomarcador en caracoles de agua dulce. Klobucar et al. (1997) demostraron que el pentaclorofenol (compuesto organoclorado, utilizado como plaguicida) inducía la peroxidación lipídica en tejido de caracoles de agua dulce (Planorbariuscorneus L.). En otros moluscos como los bivalvos se ha demostrado la inducción de peroxidación lipídica cuando se encuentran expuestos a otros xenobióticos (Singh et al., 1993; Géret et al., 2001; Ali, 2014).
Los factores fisicoquímicos y la exposición a plaguicidas podrían estar condicionando los cambios de los biomarcadores por temporada en los caracoles. En este sentido, se ha demostrado que ambos factores podrían tener una interacción importante (Pfeifer et al., 2005; Zanette et al., 2011). Diversos autores consideran que, al evaluar biomarcadores enzimáticos en campo, se deben tomar en cuenta los parámetros fisicoquímicos e incluso considerar otros factores como la disponibilidad del alimento y la masa y tamaño corporales, entre otros, debido a que estos pueden modificar la actividad de biomarcadores como la AChE y de otros asociados al estrés oxidativo en invertebrados (Sheehan y Power, 1999; Filho et al., 2001; Damiens et al., 2004; Leinio y Lehtonen, 2005; Amiard-Triquet y Berthet, 2014).
Al igual que la Ciénaga de Tamasopo, existen diversos humedales intercontinentales de importancia ecológica en México y San Luis Potosí que sufren de grandes presiones ambientales. El género Pachychilus se encuentra bien distribuido en México y Centroamérica, por lo que podría ser útil para la generación de un programa monitoreo a largo plazo para evaluar el estado de salud de los humedales. Es importante considerar complementar las mediciones de los diversos efectos biológicos con la exposición a plaguicidas (matrices ambientales y biológicas) para comprender mejor la variabilidad espacio-temporal de estos biomarcadores (Sharma y Rawat, 2009).
CONCLUSIONES
La respuesta de AChE, GST y MDA en Pachychilus sp. en diferentes temporadas en la Ciénaga de Tamasopo está relacionado con factores fisicoquímicos del agua y con la exposición a plaguicidas que se usan en los cultivos aledaños a este humedal; a su vez, estos factores modulan de manera conjunta las respuestas fisiológicas de estos organismos. Este es el primer estudio de este tipo que se realiza en la biota de este humedal, el cual podría servir como línea de base para futuras investigaciones. Este estudio demuestra la utilidad de los caracoles de agua dulce (Pachychilus sp.) como bioindicadores del estado de la salud de los ecosistemas, toda vez que estos organismos pueden encontrarse en cuerpos de agua como humedales o ríos en esta región.
Es necesario llevar a cabo una evaluación más profunda en el humedal, en la que se incluyan diferentes efectos biológicos y otros organismos de importancia ecológica (anfibios, reptiles o aves), así como la determinación de concentraciones de plaguicidas y otros contaminantes. La información conjunta seria de utilidad para definir las rutas de exposición y establecer los posibles riesgos para la salud humana y para los ecosistemas derivado de la utilización de plaguicidas en los cultivos aledaños al humedal, que deriven en medidas de gestión para su conservación y correcto manejo sustentable.

