











 Spanish (pdf)
Spanish (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mail Cited by SciELO
Cited by SciELO  Cited by Google
Cited by Google  Similars in
SciELO
Similars in
SciELO  Similars in Google
Similars in Google 
 Permalink
Permalink
INTRODUCCIÓN
El arroz es una planta monocotiledónea que pertenece al género Oryza, dentro de la familia Poaceae. Las raíces son delgadas, fibrosas y fasciculadas, el tallo consta de nudos y entrenudos, con una longitud entre los 60 y 120 cm. En cada nudo se forma una hoja y una yema la cual a su vez puede desarrollarse en un hijo o macolla (Degiovanni Beltramo et al., 2010). Las hojas son alternas con un limbo lineal y plano. Las flores están organizadas en espiguillas que forman la panícula y el ovario maduro es el grano del arroz (Bernis y Pámies, 2004). Este cultivo se puede sembrar en diferentes tipos de suelo y clima, bajo riego y en secano. El ciclo del arroz está condicionado por la temperatura, el periodo vegetativo en variedades tardías puede llegar hasta 180 días y 90 días en arroces precoces (Food and Agriculture Organization of the United Nations, [FAO], 2004). El género tiene registradas 25 especies, pero solo dos son sembradas: Oryza sativa L. y Oryzaglaberrima Steud. y se cultiva hace 8000 años (GRiSP, 2013). La especie más sembrada y con mayor distribución es O. sativa, a partir de ella se formaron tres subespecies (Índica, Japónica y Javánica) que se adaptaron a diferentes regiones (Sharma, 2010).
Entre los cereales, el arroz es el más sembrado en el mundo y suple gran parte de la demanda alimenticia. En Latinoamérica, un tercio de la energía que aprovecha la población procede de este cereal (Degiovanni Beltramo et al., 2010). El arroz forma parte de la subsistencia de unos 3000 millones de individuos, el 90 % de ellos vive en el continente asiático. Entre los países con mayor producción, en orden descendente, están: China, India, Indonesia, Bangladesh, Vietnam, Tailandia, Birmania, Filipinas y Brasil. El fitomejoramiento del arroz tiene como objetivo obtener cultivares con mayor resistencia a plagas, con un alto rendimiento y con excelente calidad molinera y culinaria; y de igual manera, que se adapten a diferentes ambientes (Torres y Martínez, 2010).
El Fondo Latinoamericano para Arroz de Riego (FLAR) realiza cruzamientos y selecciona germoplasma que luego en generaciones avanzadas (F5 a F7) es entregado a sus países miembro, en la zona tropical, por medio de viveros de observación o colecciones de las mejores líneas, llamados VIOFLAR. Una vez que los programas de fitomejoramiento de los países reciben los VIOFLAR, comienza un proceso de selección en condiciones locales que puede llevar a la selección de variedades. Los viveros que entregan el FLAR a organizaciones de sus países miembro están evaluados para todos los caracteres de interés. Estas características pueden variar a medida que los técnicos del país receptor hacen las selecciones en condiciones locales. Como consecuencia de la selección de variedades a partir de líneas avanzadas que no son 100 % homocigotas, es muy posible que la variedad liberada resulte diferente, en algunas características, de la línea que dio origen a ella.
Con el tiempo, algunas variedades, disminuyen o pierden características de interés, debido según Espe et al. (2018) a un manejo inadecuado en los procesos de fitomejoramiento o falta de adaptación a diferentes condiciones y ambientes. Uno de los factores determinantes para aumentar la producción sostenible y mantener la pureza genética de las variedades es la obtención de semilla de calidad por parte de los programas de fitomejoramiento. Esto implica que el mantenimiento varietal sea crucial para fijar las características deseadas y evitar que las variedades se degeneren con el tiempo (Lemus y Díaz, 2009). En algunos programas de fitomejoramiento se han realizado investigaciones similares para determinar la variabilidad genética en líneas, progenitores y variedades comerciales; estos aportes han permitido establecer los cambios en el tiempo que se muestran en los genotipos (Quirós, 2003). Por otra parte, Pérez-Almeida y Montoya-Arambú (2009) hicieron una evaluación de la variabilidad genética de variedades y líneas de arroz en Venezuela, donde utilizaron parámetros fenotípicos y moleculares. Los resultados mostraron que no hubo asociación entre las características moleculares y fenotípicas de las variedades y líneas. También, Reig (2019) y Aguirre et al. (2005) hicieron estudios de diversidad genética y genotipado de variedades y líneas para estudiar la distancia genética y ver aquellos caracteres con asociación. En el estudio de Aguirre et al. (2005) a través de la caracterización molecular se detectó bajo nivel de polimorfismo y diversidad genética entre las variedades y líneas; además, se pudo establecer grupos de acuerdo con la asociación de los caracteres en estudio. Los resultados mostrados por Reig (2019) permitieron diferenciar diferentes grupos de acuerdo con el origen; además, constató la asociación de los SNP con caracteres como el rendimiento y la floración.
El objetivo de este trabajo fue determinar las diferencias entre 14 variedades comerciales de arroz de América Latina y sus correspondientes líneas de origen a través de la caracterización morfo-agronómica, de calidad de grano y molecular.
MATERIALES Y METODOS
El trabajo de investigación se realizó en el Centro Internacional de Agricultura Tropical (CIAT), en Palmira, Colombia. Inicialmente, se multiplicó la semilla de las variedades para trabajar con semilla fresca y de la misma cosecha. Debido a la disponibilidad de semilla en el banco de trabajo, se estudiaron 14 variedades (suministradas por los fitomejoradores de las instituciones o empresas que las liberaron) y sus correspondientes líneas de origen FLAR (identificadas con las letras FL) (Tabla 1). Se hicieron dos siembras, en campo, con el método de trasplante y en condiciones de riego. La primera se hizo en enero y la segunda, en mayo de 2018. Las evaluaciones moleculares se hicieron a partir de ADN extraído de tejido foliar, recolectado en invernadero.
Tabla 1 Líneas origen con su correspondiente variedad, sus respectivos progenitores, el país y año de liberación de cada variedad.
| Línea origen | Variedad | Progenitores | País y año de liberación | |
|---|---|---|---|---|
| FL03187-15P-3-3P-1P-M | Variedad 1 | CT8008-16-31-3P-M/CT10308-27-3-1P-4-3-2P//FL00595-12P-1-1P-M | BOLIVIA | 2006 |
| FL03160-6P-12-2P-1P-M | Variedad 3 | ORYZICA 1/IR21015-72-3-3-3-1//CT10310-15-3-2P-4-3 | COSTA RICA | 2006 |
| FL03233-8P-6-2P-2P-M | Variedad 6 | CT10825-1-2-1-3-M/FEDEARROZ 50//FL00470-29P-7-3P-M | ECUADOR | 2009 |
| FL05372-7P-4-4P-2P-M | Variedad 4 | CT8222-7-6-2P-1X/FSR214-M-5-1-1//FL03375-3P-2-3P-4P | COSTA RICA | 2010 |
| FL04208-6P-5-2P-3P-M | Variedad 7 | CT13503-M-3-1-M-2-4P/CT8837-1-17-1P-4-M//FEDEARROZ 2000 | HONDURAS | 2010 |
| FL06885-4P-3-2P-3P-M | Variedad 8 | FL03188-7P-5-3P-3P-M-1P/CT6096-7-4-4-3-M-1P//FL03323-5P-39-3P-1P-M-1P | HONDURAS | 2010 |
| FL05372-7P-1-3P-2P-M | Variedad 11 | CT8222-7-6-2P-1X/FSR214-M-5-1-1//FL03375-3P-2-3P-4P | PANAMÁ | 2011 |
| FL07321-3P-4-3P-1P-M | Variedad 13 | SCH98-268-1-1-8-2P-2P-1P/FL001028-8P-3-2P-1P-M-8X-1P-1P-PAN1//FL03174-8P-7-2P-2P-M | PANAMÁ | 2011 |
| FL03160-6P-12-2P-1P-M | Variedad 10 | ORYZICA 1/IR21015-72-3-3-3-1//CT10310-15-3-2P-4-3 | NICARAGUA | 2012 |
| FL05482-8P-5-2P-1P-M | Variedad 14 | CT10166-16-1-2P-1-3/LV200-1-1-1-M//FL03188-7P-5-4P-M | GUYANA | 2012 |
| FL05372-7P-5-3P-1P-M | Variedad 12 | CT8222-7-6-2P-1X/FSR214-M-5-1-1//FL03375-3P-2-3P-4P | PANAMÁ | 2013 |
| FL05392-3P-12-2P-2P-M | Variedad 5 | CT9506-28-3-3P-M-1-M/FSR214-M-5-1-1//FL03191-7P-8-2P-4P | MÉXICO | 2014 |
| FL04621-2P-1-3P-3P-M | Variedad 9 | CT10825-1-2-1-3-M/FEDEARROZ 50//FL02066-4P-1-1P-M | MÉXICO | 2014 |
| FL07157-2P-4-1P-1P-M | Variedad 2 | FL04896-8P-7-3P-1P-1P//FL02066-4P-1-1P-M/FL03191-5P-10-1P-3P-M-1P | BOLIVIA | 2015 |
Evaluación fenotípica
Se utilizó un diseño de bloques completos al azar con tres repeticiones, cada parcela experimental (9,2 m2) fue de seis surcos con 17 plantas cada uno. Para las evaluaciones y toma de datos se usó como unidad experimental los cuatro surcos internos para evitar el efecto de borde. Se evaluaron 25 características morfo-agronómicas y de calidad de grano (Tabla 2). En la primera siembra, se incluyeron todas las variedades y sus líneas de origen. Luego, se hizo el análisis molecular donde se identificó la correspondencia de las variedades con su línea origen. Con base en ello, aquellas variedades que no tuvieron correspondencia se excluyeron de la segunda siembra.
Tabla 2 Características morfo agronómicas y de calidad de grano evaluadas para las variedades y sus líneas origen.
| Característica (unidad) | Descripción |
|---|---|
| Número de panículas por planta (n°) + | La cantidad de panículas totales y productivas de tres plantas por genotipo por repetición. |
| Número de macollas totales por planta (n°) | La cantidad de tallos o macollas totales de tres plantas por genotipo por repetición |
| Altura de planta (cm) + | Longitud desde la base de la planta hasta el ápice de la panícula principal. |
| Porcentaje de tallos fértiles (%) + | Número de panículas/ número total de macollas x 100. |
| Días a floración al 50 % (días) + | Número de días desde la siembra hasta cuando el 50 % de las plantas de la parcela florecieron. |
| Días a floración al 100 % (días) | Número de días hasta que todas las plantas de la parcela florecieron. |
| Longitud de panícula (cm) + | Longitud desde el nudo ciliar hasta el grano más distal de la panícula. |
| Número de granos llenos (n°) + | Número de granos llenos |
| Porcentaje de esterilidad (%) + | Número de granos vanos/ número de granos totales x 100. |
| Peso de granos por panícula (g) y Peso de mil (g) + | Peso de granos secos |
| Rendimiento (kg/planta) (kg/ hectárea) + | Peso de grano seco (12 % de humedad) por planta y por parcela. El área cosechada de la parcela fue de 5,4 m2. |
| Índice de pilada en cosecha oportuna + | Peso del grano pulido entero más el grano de % de tamaño/ el grano total de las plantas que fueron cosechadas oportunamente (humedad optima del grano entre 20 a 24 %) |
| Índice de pilada en cosecha retrasada + | Se da cuando los granos alcanzan la humedad adecuada para cosecha, y son rehidratados. Si el grano se humedece nuevamente este tiende a partirse. Se hizo el mismo procedimiento del índice de pilada en cosecha oportuna con la diferencia que los materiales fueron sometidos a procesos de simulación de cosecha retrasada según el proceso adaptado y estandarizado por el CIAT-FLAR. |
| Longitud de grano (mm) + | El largo, ancho y relación largo/ancho se determinó en mm por medio de imágenes usando el software Image J |
| Contenido de amilosa % + | Contenido de amilosa en el espectrofotómetro de infrarrojo cercano (NIRS). |
| Breakdown + | Se evaluó la viscosidad mediante el analizador rápido de viscosidad (RVA). |
| Setback + | Se evaluó la consistencia mediante el RVA. |
| Virus de la Hoja Blanca del arroz (RHBV) + y daño mecánico | En invernadero, en un diseño de bloques comple tos al azar con tres repeticiones, en dos estudios diferentes, se hicieron evaluaciones con el insecto (Tagosodes orizicolus o Sogata) vector para conocer la reacción al virus de la hoja blanca y, con el insecto no vector para conocer la reacción al daño directo causado por el insecto. |
+ Variables seleccionadas en el análisis de componentes principales.
Análisis de datos
Se hizo un análisis combinado de la varianza de datos fenotípicos de las dos siembras en campo y los respectivos contrastes ortogonales para comparar las medias de la variedad con su línea origen, respectivamente; las diferencias se establecieron con un valor p < 0,05. También se hicieron correlaciones de las variables en estudio. Para conocer las posibles relaciones entre los genotipos, usando la matriz de medias ajustadas de datos morfo-agronómicos, se hizo un análisis de componentes principales (ACP) y de conglomerados por el método de Ward o método de la varianza mínima. Para los datos de marcadores moleculares tipo SNP se calculó una matriz de distancia basada en la proporción de alelos compartidos y un análisis de conglomerados por el método de Ward.
Evaluación genotípica
La caracterización molecular se realizó a diez plantas individuales de cada material. La extracción de ADN se hizo a partir de tejido foliar. Para ello se cortó 1 cm de la parte apical de la hoja más joven de plantas con 15 días de edad, que crecieron en invernadero solo para este fin. El tejido se mantuvo en hielo hasta llegar al laboratorio donde se almacenó a -80 °C, hasta su procesamiento. La extracción del ADN se realizó siguiendo el protocolo de Risterucci et al. (2000).
La detección de los 96 marcadores tipo SNP se hizo mediante PCR-alelo específico fluorescente (tecnología SNPType de Fluidigm). Aquellos SNP con más de 10 % de datos faltantes se excluyeron del análisis para evitar imprecisiones y ambigüedades en los perfiles genéticos. Luego de aplicar este criterio, 92 SNP polimórficos permanecieron en el análisis. Se utilizaron tres patrones de referencia o controles: Nipponbare (variedad de tipo japónica templada cuyo genoma ha sido de referencia para comprender la diversidad entre miles de cultivares de arroz y sus parientes silvestres, así como los principales cultivos de cereales), IR36 e IR64 (mega variedades plantadas en gran diversidad de ambientes y usadas ampliamente en estudios científicos).
Análisis molecular
Se utilizó el programa PowerMarker v3.25 (Liu y Muse, 2005) para calcular los parámetros descriptivos de variación genética: número de loci polimórficos, loci con datos faltantes, número de genotipos observados, frecuencias alélicas, heterocigosidad y contenido de información polimórfica (PIC). Se usó el mismo programa para el análisis de agrupamiento (o de conglomerados) y para establecer las relaciones entre los individuos en estudio, con base en la distancia genética de Nei (Nei, 1972). Los dendrogramas se realizaron con el programa Mega 6 (Tamura et al., 2013) y se editaron con FigTree v.1.4.3 (Rambaut, 2016). Se utilizó el programa GenAlEx 6.502 (Peakall y Smouse, 2012) para estimar la diversidad dentro del mismo material o intra-material (Indice de Shannon) y de perfiles genéticos (Multilocus matches) para realizar el análisis de coordenadas principales y calcular el índice de diferenciación genética FST. El FST es una herramienta muy eficaz para medir el grado de diversidad presente en grupos y para corroborar el grado de diversidad que contiene un cultivar (Jakobsson et al., 2013).
Análisis combinado
Con el propósito de integrar la información genética y fenotípica se utilizó el índice de Gower para obtener la matriz de distancias de los genotipos y un análisis de conglomerados usando el método de Ward con el propósito de comprobar si existía correspondencia entre la variedad y su línea origen a partir de toda la información obtenida. Se utilizó el programa estadístico SAS® (Statistical Analysis System) versión 9,3 para Linux.
RESULTADOS Y DISCUSIÓN
Evaluación fenotípica
La capacidad de macollamiento (número de tallos en una misma planta) fue superior a 30 macollas (o tallos) por planta. El material con mayor cantidad de macollas fue la línea origen de la variedad 5, con 39. Para esta característica, en las ocho variedades y también en la variedad que coincidió con la línea hermana de su ancestro, no hubo evidencia estadística que las diferenciara; asimismo, se obtuvo un coeficiente de variación de 17,3 %. La cantidad de macollas está determinada por la radiación solar, la cantidad de agua (el déficit de agua en la etapa vegetativa limita el macollamiento), la población de las plantas y los niveles de nutrientes, entre otros factores. Un buen macollamiento hace que las plantas puedan cubrir mejor los espacios en el cultivo (Degiovanni Beltramo et al., 2004) y reducen la presencia de malezas. Esto concuerda con lo publicado por el Centro Internacional de Agricultura Tropical (CIAT, 1985) que explica que el arroz trasplantado puede desarrollar alrededor de 30 macollas y el CIAT (1983) reporta que una planta de arroz con más de 25 macollas es el tipo de planta preferido.
La media ajustada general de la altura de las líneas comparada con la de las variedades no mostró diferencias significativas y tuvo un coeficiente de variación de 5,1 %. La altura promedio de los materiales estuvo entre 90,6 y 109,3 cm; catalogados como porte bajo e intermedio. Los genotipos con estas características son deseables en un programa de fitomejoramiento (CIAT, 1985). Los valores de altura registrados en este estudio son similares a los reportados por Arce (2006). Cuando se hicieron pruebas de contrastes ortogonales para comparar la variedad y su línea origen, hubo diferencias (p = 0,00008) entre la variedad 2 (109,3 cm) y su línea origen (97,3 cm).
En cuanto al porcentaje de macollas o tallos fértiles no hubo diferencias significativas entre la línea y la variedad, los materiales estuvieron alrededor del 93 % de tallos fértiles con un coeficiente de variación de 2,7 %. Los materiales en estudio presentaron entre 28 y 37 tallos fértiles. El número de tallos fértiles es un factor importante para la obtención de buenos rendimientos y está influenciado por componentes genéticos. Se encuentran variedades con alto macollamiento, pero con bajo porcentaje de tallos fértiles (Martínez, 2010).
En la floración al 50 % yal 100 % hubo diferencias estadísticas en tres variedades y sus respectivas líneas origen, siendo las variedades 2 y 3 más precoces que sus respectivas líneas de origen, con una diferencia de hasta cinco días. Por el contrario, la variedad 14 fue tres días más tardía que su línea origen. Estas variables presentaron un coeficiente de variación de 1,4 % y 1,7 %, respectivamente. Estos cambios pueden darse por la variabilidad genética presente en los materiales, la influencia ambiental, el manejo del agua, la edad al día del trasplante, la fertilización nitrogenada, entre otras causas (CIAT, 1985; Jennings et al., 1981). Las demás variedades y líneas conservaron la característica. El período entre el inicio y final de la floración de las variedades y líneas estuvo dentro del rango de cuatro a siete días como lo menciona el CIAT (1985).
En cuanto a la longitud de la panícula, no hubo diferencias significativas, cada línea origen se comportó igual a su variedad, los materiales estuvieron entre 21,0 cm y 25,8 cm, con un coeficiente de variación de 5,7 %. La variedad 1 y su línea origen presentaron las panículas de mayor longitud con 25,5 cm y 25,8 cm, respectivamente. Estos valores son similares a los reportados por González (2015), los cuales estuvieron entre 23,2 cm y 26,3 cm. La variación reportada dentro de la subespecie índica está entre 12 cm y 40 cm (Zhang et al., 2015).
El número de granos llenos por panícula estuvo entre 110 y 174, con un coeficiente de variación de 14,8 %. Hubo diferencias significativas en la variedad 7, con 152 granos, en comparación con 121 granos en su línea origen. Entre las demás variedades y sus líneas origen, no hubo evidencia estadística que demostrara las diferencias significativas. Resultados similares han sido encontrados por diversos autores quienes mencionan que el llenado de granos es una de las características de rendimiento donde se presenta mayor variación (Golam et al., 2011; Saha et al., 2015).
Las variedades que presentaron mayor esterilidad fueron la variedad 3 y la 12 con 16,6 % y 16,8 %, respectivamente. También, sus líneas origen mostraron valores similares 19,7 % y 17,4 %, respectivamente, sin diferencias significativas con sus líneas de origen. Esta variable presentó un coeficiente de variación de 45,4 %, valores por debajo de 15 % de esterilidad son comunes en variedades tipo índica (Díaz et al., 2017; Jennings et al., 1981). La esterilidad puede ser resultado de las temperaturas que inciden sobre el polen, por debajo de los 20 °C afectan la microsporogénesis, y si son superiores a los 40 °C, pueden afectar el tubo polínico. Además, el exceso de nitrógeno, enfermedades e insectos podrían ser otros factores influyentes en esta característica (Espino, 2014). Con respecto a las variedades que mostraron valores superiores al 15 % de esterilidad, podría deberse a factores genéticos debido a que sus líneas origen presentaron valores similares.
Con respecto al peso de la panícula no hubo diferencias significativas entre la variedad y su línea origen, los valores estuvieron entre 2,8 y 3,9 g (CV= 18,1 %), la línea origen de la variedad 12 fue la que tuvo el mayor peso con 3,9 g. Las panículas pesadas son favorables para una mejora en el potencial de rendimiento, pero requieren periodos largos de floración y tiempo en el llenado de granos por lo que este carácter es muy vulnerable a cambios ambientales (Cheng et al., 2007).
En cuanto al peso de los mil granos, la variedad 14 y su línea origen tuvieron valores similares (29,9 y 30,1 g, respectivamente). Los materiales que presentaron los menores valores fueron la variedad 4 con 23,3 g y su línea origen con 22,6 g. Solo la variedad 2 y su línea de origen presentaron diferencias significativas (27,8 y 26,5 g, respectivamente). El coeficiente de variación fue de 3,6 %. Jennings et al. (1981) reportaron que variedades comerciales se encuentran en un rango de peso entre 20 y 35 g.
En el rendimiento por parcela hubo diferencias estadísticas (CV= 11,6 %) entre la variedad 2 y su línea origen con 3,0 y 2,5 kg, respectivamente. Los pesos de mayor valor los presentó la variedad 2 (3,0 kg), la variedad 5 (3,1 kg) con su línea origen (3,2 kg) y la variedad 7 (3,1 kg).
De acuerdo con el peso de grano por planta, la mayoría de los materiales estuvo por encima de los 50 g. Los menores rendimientos los obtuvo la variedad 3 (44,0 g) y su línea origen (44,2 g). La variedad 2 con 54,6 g fue la única diferente a su línea origen con 44,7 g. Las demás variedades coincidieron con su línea de origen.
La mayoría de los materiales presentó un porcentaje de grano blanco total, en cosecha oportuna, cercano al 70 %. Los mayores valores fueron los de la variedad 5 (74,5 %) y el de su línea origen (74,2 %). No se encontraron diferencias significativas para esta variable (CV= 1,5 %). Estos resultados concuerdan con lo publicado por Singh et al. (2000), quienes obtuvieron valores alrededor del 70 %. CIAT (1989) y Hardke y Siebenmorgen (2012) determinaron que del 30 % de pérdida del peso, el 20 % es cáscara y el restante es el salvado.
En el porcentaje de blanco entero en cosecha oportuna ninguna variedad fue diferente a su línea origen (CV= 2,0 %). Para esta variable, casi todas las variedades y su línea origen tuvieron porcentajes por encima del 70 %. La variedad 3 y su línea origen mostraron los porcentajes de menor valor con 67,5 y 67,6 %, respectivamente. Estos porcentajes son mayores que las exigencias del mercado cuyo valor aceptado es del 55 % (CIAT, 1989). Porcentajes por encima del 55 % reflejan un buen manejo del cultivo, correcto proceso de secado y además buenas características genéticas del cultivar (Xangsayasane et al. 2018).
Según la variable blanco total en cosecha retrasada, ocho variedades y su línea origen estuvieron por encima del 70 %. No hubo diferencias significativas (CV=2,7 %) entre las variedades y sus líneas origen. Esta característica mostró los mismos rangos que en la cosecha oportuna. Los materiales tuvieron alrededor del 30 % de pérdida luego del proceso de descascarado y pulido.
En cuanto al porcentaje de grano blanco entero en cosecha retrasada todas las variedades y su línea origen superaron al testigo susceptible PN1 (42,7 %) y ninguna variedad superó al testigo tolerante IRGA BR 409 (69,8 %). Las variedades 1, 5, 9 y 14 se comportaron como intermedias al igual que sus líneas origen. La variedad 7 y su línea origen fueron iguales al testigo tolerante. La variedad más afectada, en cuanto a esta característica, fue la variedad 3 y su línea origen. No hubo diferencias entre la línea origen y la variedad (CV=7,7 %). Cuando el grano está listo para la cosecha y se vuelve a humedecer sucede una rápida expansión en la superficie lo que resulta en fisuras. Esto se refleja en pérdidas de rendimiento en el molino porque el grano tiende a partirse (Hardke y Siebenmorgen, 2012). Según Berrio et al. (2002) la reducción del rendimiento en un material tolerante a la cosecha retrasada puede estar entre 0 y 20 %; los materiales susceptibles pueden perder hasta un 50 % o más del índice de pilada. En este estudio algunos materiales perdieron hasta un 55 % del índice de pilada. La mayoría de las variedades y sus líneas origen presentaron valores de rendimiento de grano entero cercanos al 60 % en cosecha retrasada. Estos valores son iguales o mayores que los aceptados en cosecha oportuna, lo que evidencia un gran valor del germoplasma FLAR en cuanto a la calidad molinera.
No se evidenciaron diferencias estadísticas en la longitud del grano (CV=1,8 %) entre las variedades y las de sus correspondientes líneas de origen. La variedad 14 y su línea origen tuvieron granos con 7,5 mm de longitud, es decir, tipo "extralargo". Cinco variedades y su correspondiente línea origen tuvieron grano largo y tres variedades y sus líneas origen, grano intermedio.
En la variable ancho de grano, dos variedades (3 y 12) y sus líneas origen fueron diferentes (CV=1,5 %). En las demás se mantuvo la característica. Los valores estuvieron entre 2,1 y 2,3 mm. En la relación largo-ancho seis variedades se clasificaron como de grano fino, al igual que sus líneas origen. Las variedades 9 y 12 y sus líneas origen tuvieron grano medio. La variedad 5 fue de grano fino, pero su línea origen fue de grano medio. En la relación largo-ancho las variedades mantuvieron la característica deseada por los consumidores, es decir, grano fino y medio (Torres y Martínez, 2010).
Con respecto al contenido de amilosa, no hubo diferencias estadísticas entre las variedades y sus respectivas líneas origen (CV= 6,7 %), siete variedades y sus línea origen resultaron con amilosa alta y dos variedades (7 y 2) y sus respectivas líneas origen con amilosa intermedia. CIAT (1989) dice que los consumidores latinoamericanos prefieren arroces sueltos, característica dada por un contenido intermedio a alto. También International Rice Research Institute (IRRI, 2006) explica que el contendido de amilosa es un parámetro relacionado con la firmeza y la adherencia, propiedades que inciden en las preferencias de los consumidores. La mayoría de los genotipos con contenidos de amilosa alto tienen una textura suave (Chen et al., 2017).
Hubo diferencias estadísticas entre la variedad 2 y su línea origen con relación a la variable Breakdown (CV=29,4). En la clasificación de acuerdo con la viscosidad, siete variedades y sus líneas origen resultaron ser de tipo sueltas. La variedad 7 y su línea origen se clasificaron de tipo intermedio. La variedad 2 fue de viscosidad intermedia, pero su línea origen fue de tipo suelto. Esto significa que las variedades con un Breakdown alto e intermedio poseen mayor estabilidad en los gránulos de almidón, eso se refleja en la soltura de los granos luego del enfriamiento (García, 2015).
Según la variable Setback (CV= 32,6 %), las variedades 7y2 y sus líneas origen se clasificaron como blandas. Cuatro variedades y su línea origen se clasificaron de tipo intermedio y la variedad 4 y su línea origen se clasificaron de tipo consistente. La variedad 3 difirió en esta característica y ocupó el rango intermedio, su línea origen se comportó como blanda. La variedad 12 se comportó como intermedia y su línea origen consistente. No hubo diferencias estadísticas entre la variedad y su línea origen, pero si en los rangos de clasificación. El Setback es una característica muy importante en las evaluaciones de los cereales porque está asociado con la amilosa y el Breakdown, valores superiores a 1600 cP se clasifican como consistentes, esto está sustentado por Tong et al. (2014) y Hsu et al. (2015) donde explican que valores altos de Setback determinan los arroces consistentes.
Los resultados de la variable resistencia al virus de la hoja blanca mostraron que no hubo diferencias significativas entre las variedades y sus líneas origen. El coeficiente de variación para esta variable fue de 55 % Todas las variedades conservaron su característica entre resistentes e intermedias. La variedad 14 fue la más afectada en esta característica al igual que su línea origen. Uno de los objetivos prioritarios del programa de fitomejoramiento del FLAR para la zona tropical, es la resistencia al virus de la hoja blanca. Las líneas que son enviadas a los países miembros presentan porcentajes altos e intermedios de resistencia. En este estudio los niveles de resistencia se han mantenido en todas las variedades, eso muestra el buen cuidado que han tenido los fitomejoradores para esta enfermedad. La variedad 14 fue susceptible al igual que su línea origen. En algunos casos la resistencia se ve influenciada por el ambiente y genera cambios en genotipos con alta resistencia (Vale et al., 2001). Ningún material evaluado fue inmune al virus como lo dijo Zeigler et al. (1988).
El daño mecánico causado por sogata (Tagosodes orizicolus Müir) a las variedades 2 y 12 y a sus líneas origen, fue consistente a través de las evaluaciones y su reacción fue de tolerancia. La variedad 5 al igual que su línea origen en ambos semestres se comportó como susceptible. En algunas variedades y líneas, los grados de daño mecánico fluctuaron entre intermedios y tolerantes en los diferentes semestres, de acuerdo con Jennings et al. (1981) la herencia para la reacción al daño mecánico se debe a un gen dominante que confiere tolerancia, pero que está condicionado a diferentes factores como a un gen modificador que confiere menor o mayor resistencia y está determinado por el genotipo, la presión y el tiempo de exposición de la planta al ataque del insecto.
Correlaciones fenotípicas
La altura y la longitud de la panícula tuvieron un valor de correlación bajo, pero significativo (0,30) lo que indica que hay una tendencia a que, a mayor altura de planta, mayor tamaño de la panícula. Esto concuerda con las evaluaciones de Díaz et al. (2000) y Villao (2017) quienes encontraron correlaciones similares.
La longitud de la panícula se correlacionó con el peso de mil granos (0,47) y el largo de grano (0,65), esto determina que a mayor longitud de panícula hubo mayor peso de mil granos y granos más largos. En los trabajos de Díaz et al. (2000) y Amela et al. (2008) también encontraron correlaciones positivas entre estas variables.
El peso de mil granos se correlacionó positivamente con el largo de grano (0,68) y negativamente con las variables de calidad molinera como blanco entero en cosecha oportuna (-0,45). Es decir, que entre más largo es el grano habrá mayor peso de los mil granos, pero menos rendimiento molinero, esto también fue reportado por Sandhu et al. (2018).
El rendimiento de la parcela tuvo una alta correlación con el rendimiento por planta (0,91). Esto indica que, en estas condiciones, la estimación de rendimiento pudo hacerse por cualquiera de las dos variables. También tuvo una asociación significativa con la variable blanco total en cosecha retrasada (0,37). Además, a mayor rendimiento se obtuvo mayor blanco total en cosecha retrasada. Lo contrario reportó Amézquita (2012) al correlacionar la variable rendimiento (al 14 % de humedad) con las variables de rendimiento molinero.
El rendimiento de blanco total en cosecha oportuna se correlacionó con el blanco total en cosecha retrasada (0,84) como lo reportado por Amézquita (2012).
El rendimiento de blanco entero en cosecha oportuna estuvo correlacionado con el blanco total en cosecha retrasada (0,81) y con el blanco entero en cosecha retrasada (0,52). Por otra parte, el blanco entero en cosecha oportuna se asoció negativamente con el largo de grano (-0,41), Sandhu et al. (2018) reportaron pérdida de rendimiento en las variedades que evaluaron debido a la mayor longitud del grano.
El contenido de amilosa se correlacionó negativamente con el Breakdown (-0,43) y positivamente con el Setback (0,65). A mayor contenido amilosa se tendrá menor Breakdown, valores bajos de Breakdown son característicos de arroces sueltos, valores altos de Setback son característicos de arroces duros. Esto concuerda con Tong et al. (2014) y Hsu et al. (2015) quienes reportaron este tipo de asociaciones.
Análisis de componentes principales
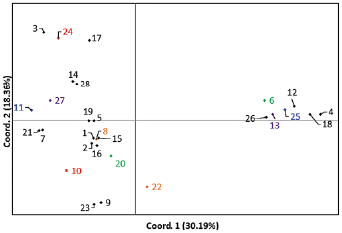
Para llevar a cabo el agrupamiento se tomaron en cuenta 17 variables cuantitativas de las 25 en estudio, esto se hizo con el propósito de reducir la dimensionalidad de los datos y al mismo tiempo tener una mejor respuesta de los descriptores en estudio. Las variedades se agruparon de la misma forma como en el análisis genotípico. Las ocho variedades se agruparon con su línea de origen y una variedad se agrupó con la línea hermana de su ancestro (Fig. 2). Los valores propios, la frecuencia absoluta y acumulada muestran el aporte en cada componente principal en este caso con cinco componentes se explica el 87,26 % de la variabilidad total. El primer componente explica el 30,5 % de la variabilidad, el segundo explica el 20,94 %, el tercero, 15,65 %, el cuarto y quinto 11,8 % y 8,38 %, respectivamente. El primer componente estuvo representado por las variables de calidad molinera como blanco entero en cosecha oportuna y retrasada, la longitud de panícula, el número de granos llenos, el peso de mil granos, el largo de grano y la variable estimada de t ha-1. El segundo componente estuvo caracterizado por las variables de calidad culinaria como amilosa, Breakdown y Setback. En el tercer componente se agruparon las variables agronómicas como número de panículas, tallos fértiles, floración al 50 % y esterilidad. El cuarto componente lo explica la variable resistencia al virus de la hoja blanca (RHBV). El quinto componente lo explican las variables altura y ancho de grano.
Evaluación genotípica
El genotipado fue exitoso para un 95,84 % de los marcadores. El análisis dentro de cada material o intra-material (Tabla 3) mostró niveles altos de heterocigosidad en algunas variedades y líneas los cuales indican grados de impureza genética y advierten problemas para los multiplicadores de semilla (Cortés-olmos et al. 2015). Esta variabilidad podría deberse a la mezcla de semillas por errores en los procesos o a la contaminación con polen indeseado, factores como estos pueden llevar a que una variedad cambie significativamente sus características a través del tiempo y no coincida con su línea origen (Casals et al., 2011). Aunque en el arroz los niveles de polinización cruzada son bajos, en algunas condiciones puede ser hasta 5 %, esto puede conllevar a que con el tiempo se den cambios en las variedades (Vallejo y Estrada, 2013). También, podría suponerse que cruzamientos espontáneos sucedieron en estas variedades.
Tabla 3 Índice de diversidad intra-material, heterocigosidad y número de perfiles genéticos estudiados en diez plantas por variedad y línea de origen FLAR.
| No | Material | Shannon (I) | Heterocigosidad (H) | No. perfiles genéticos observados | Plantas atípicas detectadas |
|---|---|---|---|---|---|
| 1 | Variedad 1 | 0,061 | 0,035 | 3 | 1 |
| 2 | Variedad 2 | 0,183 | 0,129 | 10 | 0 |
| 3 | Variedad 3 | 0,023 | 0,013 | 2 | 1 |
| 4 | Variedad 4 | 0,004 | 0,002 | 2 | 0 |
| 5 | Variedad 5 | 0,103 | 0,067 | 9 | 1 |
| 6 | Variedad 6 | 0,051 | 0,036 | 3 | 0 |
| 7 | Variedad 7 | 0,025 | 0,017 | 5 | 0 |
| 8 | Variedad 8 | 0,081 | 0,048 | 5 | 1 |
| 9 | Variedad 9 | 0,008 | 0,006 | 2 | 2 |
| 10 | Variedad 10 | 0,000 | 0,000 | 2 | 0 |
| 11 | Variedad 11 | 0,046 | 0,032 | 6 | 0 |
| 12 | Variedad 12 | 0,139 | 0,093 | 4 | 0 |
| 13 | Variedad 13 | 0,064 | 0,045 | 6 | 0 |
| 14 | Variedad 14 | 0,004 | 0,002 | 2 | 0 |
| 15 | FL03187-15P-3-3P-1P-M | 0,021 | 0,016 | 8 | 0 |
| 16 | FL07157-2P-4-1P-1P-M | 0,211 | 0,155 | 10 | 0 |
| 17 | FL03160-6P-12-2P-1P-M | 0,232 | 0,145 | 10 | 1 |
| 18 | FL05372-7P-4-4P-2P-M | 0,130 | 0,081 | 10 | 1 |
| 19 | FL05392-3P-12-2P-2P-M | 0,071 | 0,053 | 10 | 0 |
| 20 | FL03233-8P-6-2P-2P-M | 0,054 | 0,039 | 9 | 0 |
| 21 | FL04208-6P-5-2P-3P-M | 0,039 | 0,027 | 9 | 0 |
| 22 | FL06885-4P-3-2P-3P-M | 0,022 | 0,017 | 8 | 0 |
| 23 | FL04621-2P-1-3P-3P-M | 0,021 | 0,016 | 5 | 0 |
| 24 | FL03160-6P-12-2P-1P-M | 0,064 | 0,048 | 10 | 0 |
| 25 | FL05372-7P-1-3P-2P-M | 0,057 | 0,041 | 7 | 0 |
| 26 | FL05372-7P-5-3P-1P-M | 0,191 | 0,116 | 10 | 1 |
| 27 | FL07321-3P-4-3P-1P-M | 0,028 | 0,021 | 8 | 0 |
| 28 | FL05482-8P-5-2P-1P-M | 0,028 | 0,020 | 9 | 0 |
Los SNP permitieron determinar varios grupos genéticos dentro del germoplasma evaluado. Ocho de las 14 variedades se agruparon con su ancestro FL (Fig. 1). Para la variedad 12 los individuos estuvieron más cercanos a la línea hermana FL05372-7P-1-3P-2P-M, que a su ancestro o línea de origen FL05372-7P-5-3P-1P-M. Las variedades que coincidieron con su ancestro estuvieron en su mayoría en un nivel moderado de diferenciación según el coeficiente FST (entre 0,05 y 0,15) y las que no correspondieron entraron en la categoría de diferenciación muy alta (>0,25) (Tabla 4).
Tabla 4 Comparación variedad-línea origen por medio del índice de diferenciación genética
| Variedad | Línea origen | Valor FST | Nivel de diferenciación |
|---|---|---|---|
| Variedad 1 | FL03187-15P-3-3P-1P-M | 0,052 | Moderado |
| Variedad 2 | FL07157-2P-4-1P-1P-M | 0,110 | Moderado |
| Variedad 3 | FL03160-6P-12-2P-1P-M | 0,104 | Moderado |
| Variedad 4 | FL05372-7P-4-4P-2P-M | 0,107 | Moderado |
| Variedad 5 | FL05392-3P-12-2P-2P-M | 0,051 | Moderado |
| Variedad 6 | FL03233-8P-6-2P-2P-M | 0,820 | Muy alto |
| Variedad 7 | FL04208-6P-5-2P-3P-M | 0,051 | Moderado |
| Variedad 8 | FL06885-4P-3-2P-3P-M | 0,611 | Muy alto |
| Variedad 9 | FL04621-2P-1-3P-3P-M | 0,244 | Alto |
| Variedad 10 | FL03160-6P-12-2P-1P-M | 0,877 | Muy alto |
| Variedad 11 | FL05372-7P-1-3P-2P-M | 0,835 | Muy alto |
| Variedad 12 | FL05372-7P-5-3P-1P-M | 0,286 | Muy alto |
| Variedad 13 | FL07321-3P-4-3P-1P-M | 0,815 | Muy alto |
| Variedad 14 | FL05482-8P-5-2P-1P-M | 0,324 | Muy alto |
*Bajo: 0,0-0,050; moderado 0,050-0,15; alto 0,15-0,25; muy alto: >0,25 muy alto
En un estudio similar Wang et al. (2014) obtuvieron valores de FST entre 0,036 a 0,164 lo cual les permitió clasificar la mayoría de accesiones con diferenciación baja.
Para las variedades que no coincidieron con su línea de origen, se recomienda realizar una depuración mediante la eliminación de genotipos que no corresponden a la variedad (Caramante et al., 2009) y así recuperar la uniformidad intra-material. Para la estabilización genética de las variedades, la herramienta molecular juega un papel importante para reducir el trabajo en la identificación de aquellos materiales contaminados e identificar el genotipo original de la variedad (Cortés-Olmos et al., 2015). Dada la no coincidencia en algunas variedades con su línea origen, el programa de fitomejoramiento del FLAR y los programas de los países miembro, se deben establecer procesos que aseguren el mantenimiento de la pureza genética de la semilla. Además, establecer la huella genética de estas variedades y así, mantener las características de los cultivares seleccionados.
A partir de estos resultados moleculares y basados en los análisis de los datos agronómicos de la primera siembra, también se observó asociación entre las ocho variedades y sus líneas de origen en ambos análisis, de la misma manera, sucedió con la variedad que coincidió con la línea hermana de su ancestro; se excluyeron del análisis general en la segunda siembra, las variedades que no coincidieron con su línea origen (Variedades 6, 8, 10, 11, 12, 13) (Fig. 1).
Combinación de datos morfo-agronómicos, de calidad de grano y moleculares
El dendrograma establecido a partir de características morfo-agronómicas, de calidad y moleculares agrupó las variedades con sus respectivas líneas de origen (Fig. 3). En las nueve variedades que coincidieron en el análisis tanto molecular como fenotípico también coincidieron cuando se analizaron en conjunto y usando el coeficiente de Gower. Los valores reflejan que la variedad 1 con su línea origen tuvieron la mayor similitud (98 %). La variedad 2 con su línea origen fueron las de menor similitud (80 %).

Figura 2 Dendrograma establecido por varianza mínima de Ward y basado en características morfo-agronómicas de variedades de arroz y sus respectivas líneas origen.
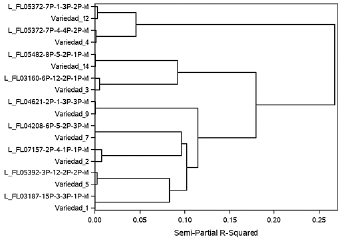
Figura 3 Dendrograma establecido por características moleculares y morfo-agronómicas de las variedades y sus respectivas líneas de origen FLAR.
Según Chávez Esponda et al. (2010) el coeficiente de Gower juega un papel importante y da mayor confiabilidad a los resultados cuando se tienen datos morfológicos y moleculares. Además, Correa Londoño et al. (2007) explican que en su estudio fue ventajoso utilizar el coeficiente de Gower para poder detectar, de una mejor manera la variabilidad. Esta investigación difiere con lo realizado por Quirós (2003) quien no encontró asociación entre los datos morfológicos y moleculares debido a la variación en las características cuantitativas. Resultados encontrados por Tapia (1998) muestran que la combinación de los datos morfológicos y moleculares fueron significativos y coherentes en la verificación de la variabilidad.
CONCLUSIONES
El uso de descriptores morfo-agronómicos permitió determinar las diferencias fenotípicas de las 14 variedades y sus líneas de origen FLAR. La variabilidad observada mediante la evaluación molecular permitió establecer el nivel de diversidad genética de las variedades y sus líneas de origen y por medio de esto se pudo esclarecer la correspondencia molecular entre ambas. De las 14 variedades en estudio, ocho coincidieron con su línea de origen. Hubo una variedad que se agrupó con una línea hermana de su ancestro. Tanto la heterogeneidad genética entre las variedades y sus líneas de origen, como la falta de homocigosis completa de las variedades sugieren que los programas de fitomejoramiento deben realizar la pronta purificación de la semilla y el establecimiento de una huella genética. Los resultados obtenidos a través del análisis morfo-agronómico y molecular, realizados tanto en forma conjunta como independiente, fueron consistentes en el agrupamiento de las variedades con sus respectivas líneas de origen.

