Spanish (pdf)
Spanish (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mail Cited by SciELO
Cited by SciELO  Cited by Google
Cited by Google  Similars in
SciELO
Similars in
SciELO  Similars in Google
Similars in Google 
 Permalink
Permalink
Caiman crocodilus Linnaeus, 1758 es el único aligatórido distribuido desde México hasta Brasil (Medem, 1983). Su rango de distribución ha derivado en una gran variación morfológica y en el reconocimiento de cuatro subespecies. A nivel del cráneo es posible encontrar dos grupos, las formas longirostrinas (cráneo con hocico angosto), con distribución cisandina, como C. c. crocodilus distribuida en Colombia, Venezuela y Brasil (Medem, 1981; 1983; Rueda Almonacid et al., 2007), y C. c. apaporiensis con una distribución restringida al Departamento de Vaupés en Colombia (Medem, 1955); por otro lado, las formas brevirostrinas (cráneo con hocico ancho) y transandinas, como C. c. chiapasius distribuida en México, Guatemala y norte del Salvador (Venegas-Anaya et al., 2008); y C. c. fuscus que se distribuye desde El Salvador hasta Ecuador (Medem, 1981; Rueda-Almonacid et al., 2007).
La variación ontogenética en la forma del cráneo de C. crocodilus, durante el desarrollo postembrionario, fue analizada por Angulo-Bedoya et al. (2019). Estos autores encontraron un patrón de desarrollo ontogenético único para C. c. apaporiensis; además encontraron diferencias entre las subespecies cis y transandinas. Finalmente, Falcón-Espitia y Jerez (2021) analizaron la variación ontogenética de tres caracteres craneales para las subespecies de C. crocodilus en Colombia, y reportaron diferencias de utilidad taxonómica.
Por lo tanto, los análisis del desarrollo craneal en C. crocodilus constituyen una fuente de información taxonómica, pero también ecológica ya que en Crocodilia el cráneo exhibe variaciones asociadas con la dieta, relacionadas con cambios en la forma del cráneo y el desarrollo de la musculatura mandibular que impactan en el uso del hábitat de estas especies durante la ontogenia (van Drongelen y Dullemeijer, 1982; Fernández-Blanco et al., 2018). Por lo tanto, el objetivo del presente estudio fue determinar el patrón de desarrollo del cráneo en C. c. fuscus durante la ontogenia postnatal en C. c. fuscus, con base en la variación de la forma craneal y de los elementos óseos que lo componen.
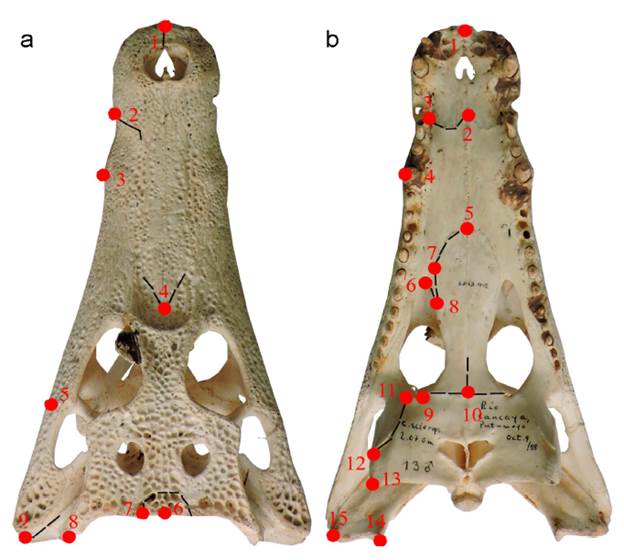
Se analizó una serie ontogenética posnatal de cráneos de C. c. fuscus de Colombia, depositados en la Colección de Reptiles del Instituto de Ciencias Naturales (ICN-UNAL, Colombia). Los cráneos se clasificaron con base en la longitud total de los individuos (LT; RBE, 2010): crías (4180 cm; n= 7); juveniles (81-120 cm; n= 3); subadultos (121160 cm; n= 9) y adultos (más de 161 cm; n= 11). El análisis de morfometría geométrica se realizó con fotografías de los cráneos en vista dorsal y ventral en el software MorphoJ (Klingenberg, 2011). Se digitalizaron nueve landmarks en vista dorsal y 15 en ventral (Material suplementario 1) en TPSDig2 (Rohlf, 2017) solo en el lado izquierdo, siguiendo la propuesta de Falcón-Espitia y Jerez (2021). Tras realizar un análisis generalizado de Procrustes, se exploró la forma craneana de las diferentes clases de edad a través de un Análisis de Componentes Principales (ACP). Luego se analizó la relación tamaño-forma (alometría) a través de una regresión múltiple de las coordenadas de Procrustes de cada uno de los landmarks contra el logaritmo del tamaño centroide y se aplicó una prueba de permutaciones con 10000 repeticiones (Drake y Klingenberg, 2008). Los residuales de la regresión se utilizaron para definir el morfoespacio de las clases de edad a través de un ACP. Finalmente, se describió la variación de los elementos craneales por observación directa de los elementos óseos entre los grupos de edad, con base en Romer (1956).
En el ACP efectuado en la vista dorsal del cráneo, no corregido por tamaño, los primeros dos componentes principales (CPs) explicaron el 73,02 % de la variación, mientras que en vista ventral explicaron el 56,19 % de la variación. Al remover el efecto del tamaño, utilizando los residuales de la regresión para redefinir el morfoespacio craneal en un nuevo ACP, la variación explicada por los primeros dos CPs en vista dorsal disminuyó al 65,31 %; mientras que, en vista ventral, los dos primeros CPs explicaron el 54,82 % de la variación. Así, el efecto calculado de la alometría sobre la muestra fue de 26,4 % (p = 0,0001; Fig. 1a) para la vista dorsal y 10,7 % (p = 0,0093; Fig. 1b) para la vista ventral, mostrando una fuerte asociación entre la forma y el tamaño en el cráneo de C. c. fuscus.
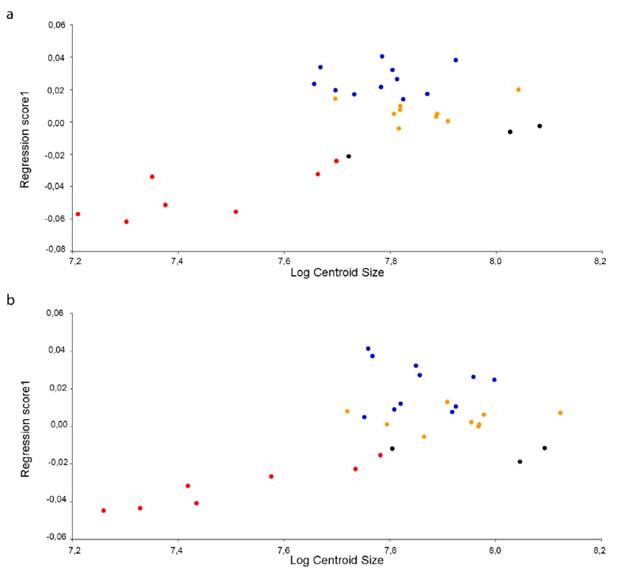
Figura 1 Regresión de las coordenadas de Procrustes vs Log del tamaño centroide en individuos de diferentes clases de edad de Caiman crocodilus fuscus. El eje X muestra el cambio en el tamaño, mientras el eje Y muestra el cambio alométrico de la forma (Regression score). (a) Vista dorsal; (b) Vista ventral. Crías (Rojo), Juveniles (Negro), Subadultos (Naranja), Adultos (Azul).
Una vez retirado el efecto del tamaño, los grupos de edad se separaron en el morfoespacio (Fig. 2). Las crías y juveniles presentaron hocicos cortos y angostos (longirostrinos), que se alargan y ensanchan durante el desarrollo ontogenético, desplazándose desde los valores negativos hacia los positivos del CP1. Los subadultos y adultos mostraron cráneos anchos y robustos, típicos del cráneo brevirostrino en C. c. fuscus, especialmente para la vista ventral (Fig. 2b). Además, respecto a las crías y juveniles, se registraron cambios de algunos elementos óseos en subadultos y adultos: engrosamiento de la base del escamoso y el cuadratoyugal; el proceso lateral del pterigoides y el proceso retroarticular del articular se hicieron más robustos; además, se desarrolló una cresta dorsal en el hueso surangular.
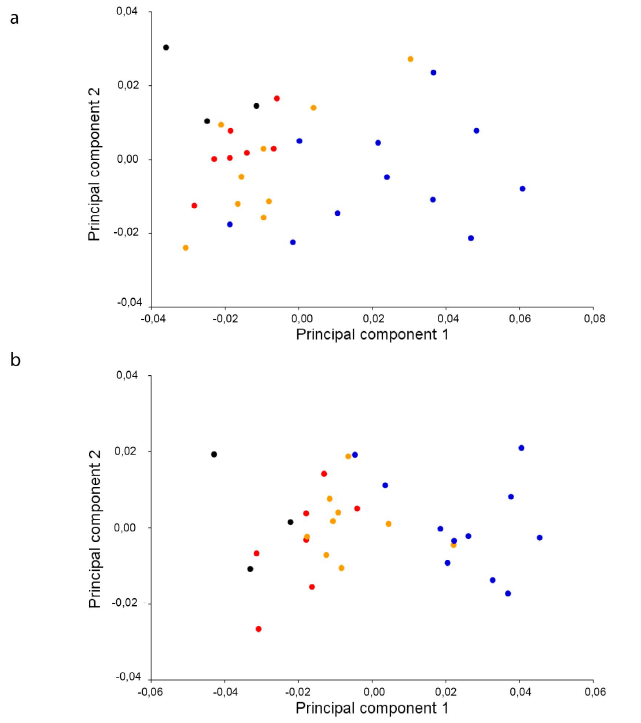
Figura 2 Análisis de Componentes Principales realizado con los residuales derivados de la regresión del tamaño sobre la forma craneal en Caiman crocodilus fuscus. (a) Vista dorsal; (b) Vista ventral. Crías (Rojo), Juveniles (Negro), Subadultos (Naranja), Adultos (Azul).
Por lo tanto, en C. c. fuscus se registraron cambios generales en la forma del cráneo durante la ontogenia postembrionaria, desde un cráneo longirostrino en neonatos a un cráneo brevirostrino en adultos, bajo un efecto alométrico estadísticamente significativo y, además, con variaciones específicas en algunos elementos óseos asociados a la región mandibular del cráneo.
En cocodrilianos se han descrito cambios morfológicos abruptos en la transición cría-juvenil (Hall y Portier, 1994). Específicamente para C. crocodilus,Ayarzagüena (1984) describió un rápido alargamiento del hocico en esta transición, tras lo cual el cráneo se robustece progresivamente en la etapa de subadulto; en el caso de C. c. fuscus analizado aquí, el mismo patrón de crecimiento fue encontrado (Figs. 1 y 2). Este patrón de variación en la forma del cráneo fue descrito para otras especies del género Caiman (Fernández-Blanco et al., 2018; Angulo-Bedoya et al., 2019) y podría ser útil en la discriminación de especies de cocodrílidos. Si bien algunos autores destacan que no es útil a nivel de subespecies (Watanabe y Slice, 2014), el análisis de Angulo-Bedoya et al. (2019) demostró diferencias en el patrón de desarrollo del cráneo entre las subespecies de Colombia, no solo entre aquellas con distribución cis y transandina, sino además destacan el patrón particular de C. c. apaporiensis. Información que, unificada a las diferencias a nivel de los elementos craneales entre subespecies, reportadas por Falcón-Espitia yJerez (2021), no solo aportan al diagnóstico de estas subespecies, sino a futuros estudios taxonómicos que reconsideren la nominación de estas entidades.
El cambio desde una forma longirostrina hacia una forma brevirostrina en C. c. fuscus puede relacionarse con la variación de la dieta de esta subespecie durante la ontogenia. Las crías y juveniles se alimentan principalmente de presas blandas, como artrópodos, mientras que los subadultos y adultos basan su dieta en el consumo de presas más grandes y duras, como vertebrados (Ayarzagüena, 1984; Bontemps et al., 2016). Este patrón ontogenético y su relación con la dieta es conocido en otros caimaninos, con implicaciones taxonómicas, funcionales y ecológicas en Caiman latirostris y C. yacare (Fernández-Blanco et al., 2018).
Es así como, en los estados de subadulto y adulto se observaron cambios en elementos óseos claves en el desarrollo de músculos que favorecen la fuerza de mordida. Por lo tanto, el ensanchamiento de la base del escamoso y el cuadratoyugal, además del proceso retroarticular del hueso articular y los procesos laterales del pterigoides otorgan un área de inserción mayor para músculos de tipo aductor, como los pterigoideos, lo que aumenta la fuerza de mordida en adultos (van Drongelen y Dullemeijer, 1982; Cleuren et al., 1995; Monteiro et al., 1997). Estas características se relacionan con funciones biomecánicas que impactan la dieta, ya que una morfología craneana específica se relaciona con hábitos alimenticios particulares en los cocodrílidos (Pierce et al., 2008). El hecho de que C. c. fuscus se alimente de presas duras permite diferenciarlo morfológica y ecológicamente de las subespecies longirostrinas como C. c. apaporiensis, que presenta hábitos mayormente piscívoros, asociados con su morfología craneana (Ayarzagüena, 1984).
En C. c. fuscus, la forma brevirostrina es el resultado de un efecto alométrico durante el desarrollo postembrionario y se relaciona con cambios en huesos del cráneo que permiten mayor fuerza de mordida en los adultos. Conocer los factores asociados a la ecología trófica de estas subespecies, en relación con los cambios craneales, es fundamental para comprender su evolución y en el establecimiento de políticas de conservación para cada una de las subespecies de C. crocodilus.