Spanish (pdf)
Spanish (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mail Cited by SciELO
Cited by SciELO  Cited by Google
Cited by Google  Similars in
SciELO
Similars in
SciELO  Similars in Google
Similars in Google 
 Permalink
Permalink
INTRODUCCIÓN
A través de las redes tróficas se puede representar los flujos de energía transferidos mediante el consumo por medio de los niveles tróficos y se puede visualizar mediante la esquematización de las relaciones de consumo (Giacomini y Petrere, 2010). A partir de dichas relaciones se pueden calcular sus propiedades que permiten conocer la estabilidad, complejidad y densidad de vínculos (De Ruiter et al. 2005) de los organismos que habitan en dicho ecosistema, basados en los análisis de los nodos, eslabones y niveles tróficos (Andramunio y Caraballo, 2012). De esta manera, el análisis de las redes permite entender la estructura y funcionamiento de los ecosistemas, ayudando a obtener un entendimiento de la composición de las comunidades que lo componen y la transferencia energética (Tamara et al. 2019).
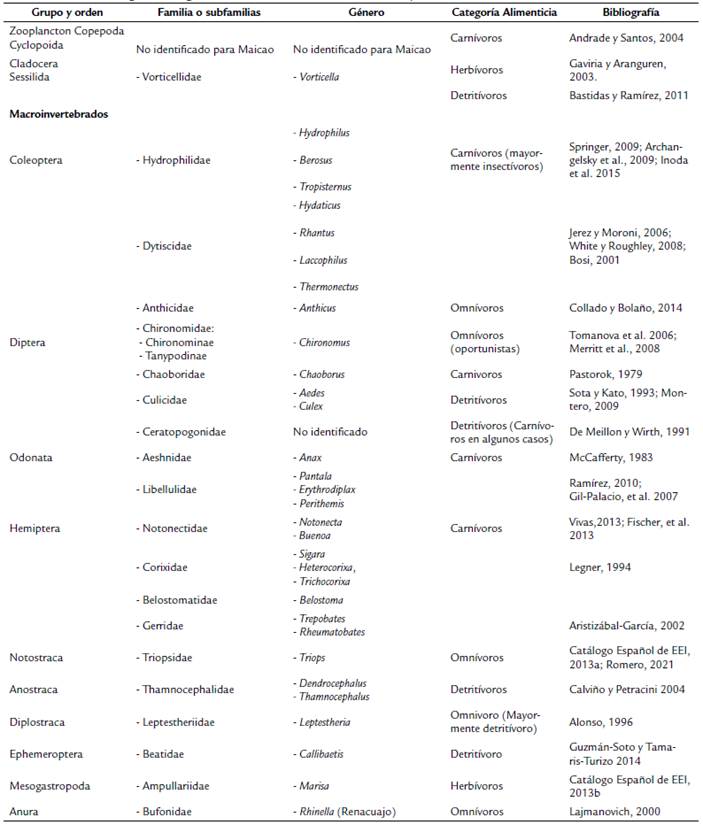
Los macroinvertebrados acuáticos desempeñan un rol importante en la transferencia de energía dentro de las redes tróficas de ecosistemas acuáticos (Allan y Castillo, 2007) comprendiendo una gran parte de la diversidad en los sistemas dulceacuícolas (Ríos et al., 2013), los cuales son dominados por insectos y crustáceos en abundancia y diversidad (Almonacid y Molinero, 2012) con depredadores dominantes como los Hemiptera, Odonata y Coleoptera (Serna-Macías et al. 2019), por ello, la importancia de entender su relaciones alimenticias.
Las charcas estacionales son ecosistemas lénticos dulceacuícolas con amplias variaciones en su tamaño, profundidad y temporalidad; se inundan durante los periodos de lluvias, al cual le sigue un período de desecación hasta que las lluvias inunden el sitio nuevamente (Keeley y Zedler, 1998). Estas variaciones ocurren como consecuencia de la dinámica de las precipitaciones locales (Williams, 2006).
En Colombia solo se conocen estudios asociados a las charcas estacionales en la región Caribe, al norte del departamento de la Guajira, enfocados en aspectos taxonómicos de organismos zooplanctónicos (Cohen et al. 2014; Fuentes-Reinés et al. 2019; Fuentes-Reinés et al. 2021), así como estudios de caracterización biológica de grupos de invertebrados y vertebrados en cinco charcas temporales de los municipios de Maicao, Manaure, Uribia, Mayapo y El Ebanal (Tamaris-Turizo et al. 2020). Lo anterior deja en evidencia que son ecosistemas poco conocidos, por tanto, la naturaleza del trabajo es exploratoria. El objetivo de este estudio fue conocer las propiedades de las redes tróficas de dos charcas estacionales en el norte del departamento de la Guajira, Colombia con base en las comunidades de macroinvertebrados y vertebrados acuáticos a través de análisis de contenidos estomacales.
MATERIALES Y MÉTODOS
Área de estudio
El departamento de La Guajira se encuentra localizado en el norte de Colombia, se caracteriza por tener un clima semiárido, suelos con formaciones aluviales y lacustres por debajo de los 100 m de altitud (CorpoGuajira, 2009). Cuenta con una vegetación donde predominan los bosques subxerofíticos con pequeño porte (Halffter y Ezcurra, 1992) y un régimen de lluvias bimodal con precipitaciones entre 200 y 1500 mm anuales (Orjuela-Rojas et al. 2011).
En este departamento se presentan condiciones acentuadas de aridez durante la mayor parte del año por la acción de los vientos alisios y la poca precipitación (IDEAM, 2014). No obstante, durante los periodos de lluvias se forman charcas estacionales que pueden albergar una alta diversidad de organismos (Fuentes-Reinés et al. 2019; Tamaris-Turizo et al. 2020).
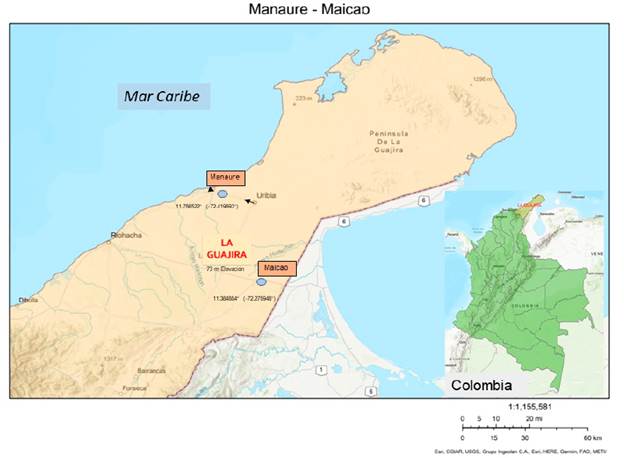
Se seleccionaron dos charcas estacionales, una en el municipio de Manaure y otra en el de Maicao (Suplemento 1), teniendo en cuenta las condiciones de seguridad y accesibilidad a zonas de muestreo. Estos ecosistemas están sujetos a disturbios antrópicos como los derivados de la fertilización de suelos agrícolas y ganaderos de zonas cercanas (López, 2014), además de disposición de desechos a causa de la cercanía de las carreteras, centros urbanos y rancherías (pequeñas poblaciones de indígenas Wayú).
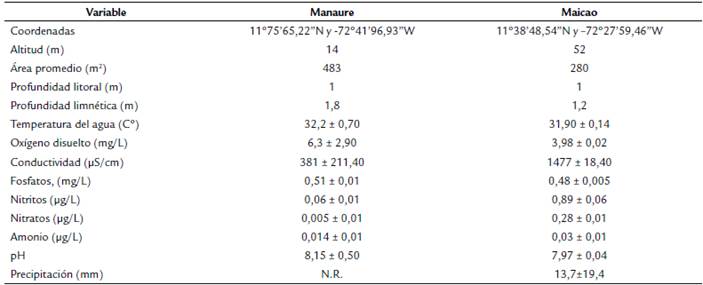
La charca Manaure posee poca vegetación en sus alrededores, con dominio de plantas de las familias Fabaceae y Cactaceae. La charca Maicao se localiza en la entrada al municipio que lleva el mismo nombre y presentó vegetación ribereña dominada por las familias Fabaceae, Cactaceae, Araceae y Onagraceae. Las charcas difirieron en el tamaño del espejo de agua, cercanía a la carretera y profundidad (Tabla 1).
Recolecta e identificación de muestras
Durante los dos muestreos se midieron las variables físicas y químicas con un medidor portátil multiparámetro (OHAUS ST-300): temperatura del agua (°C), pH, conductividad (µS/cm) y oxígeno disuelto (mg/L). Adicionalmente, se tomaron muestras de agua con botellas de 500 mL y se mantuvieron refrigeradas hasta su transporte al Laboratorio de Calidad del Agua de la Universidad del Magdalena con el fin de analizar las concentraciones de nitratos (µg/L) y nitritos (µg/L), amonio (µg/L) y fosfatos (mg/L), siguiendo métodos estandarizados (Apha, 1998).
En cada charca se realizaron dos muestreos durante el periodo de aguas altas: en noviembre del 2018 y enero del 2019 en la charca de Manaure y en octubre y noviembre del 2018 en la de Maicao. Los organismos acuáticos se recolectaron realizando arrastres en la zona litoral y limnética durante treinta minutos con una red tipo D-net de 250 µm de abertura de malla. Los organismos se almacenaron en bolsas herméticas y en frascos plásticos con etanol al 95 % y se trasladaron al laboratorio para su posterior identificación.
En el laboratorio las muestras se separaron usando tamices de 250 µm de apertura de malla, luego se depositaron en bandejas para hacer la respectiva separación. Finalmente, los organismos se preservaron en viales rotulados con etanol al 70 %. Los especímenes se identificaron al nivel taxonómico más detallado posible (nivel de genero para la mayoría de los casos), empleando las claves taxonómicas y guías de Epler (2001), Merritt et al. (2008) y Domínguez y Fernández (2009).
Análisis trófico
Una vez identificados los especímenes, se procedió a medirlos y se les extrajeron los estómagos para analizar sus contenidos mediante la técnica propuesta por Tomanova et al. (2006), con las modificaciones realizadas por Tamaris-Turizo et al. (2018). Se extrajeron estómagos de dos a 25 individuos por cada taxón y se montaron en láminas portaobjetos semipermanentes con glicerina, para analizar 20 campos por lámina, usando objetivos de 10X, 20X y 40X.
Los organismos que presentaron bajas abundancias (menor a dos individuos) no se diseccionaron porque un bajo número de muestras puede afectar la calidad de los análisis. La información sobre sus hábitos alimenticios se complementó mediante una revisión de literatura, la cual se aplicó a seis taxones de la charca de Manaure debido a la baja abundancia de individuos (menor o igual a dos:
Triops, Aedes, larva de Rhantus, Berosus, Anthicus y Anax). Además de ocho taxones pertenecientes al orden Hemiptera (suborden heteróptera), los cuales no se diseccionaron porque se alimentan mediante la succión de fluidos de vegetales y animales (Vivas y López, 2013), por ello, en sus estómagos solo se encontrarían fluidos que no se pueden identificar: Notonecta, Buenoa, Belostoma, Sigara, Heterocorixa, Trichocorixa, Trepobates y Rheumatobates. Asimismo, no se diseccionaron cuatro taxones de la charca de Maicao debido a la baja abundancia de individuos: Chironomus, Hydrophilus, Perithemis, y Pantala. Con base a los contenidos estomacales y la revisión bibliográfica se establecieron categorías tróficas para todos los taxones identificados (Suplemento 2).
Análisis de los datos
Se realizó un análisis de diversidad ( q D) basado en la serie de números de Hill evaluando los números efectivos de los taxones (Jost, 2006) para los siguientes órdenes q=0, q=1 y q=2. En los números de Hill, q=0 cuantifica la riqueza total de taxones, q=1 revela el número de taxones con similares abundancias mediante el cálculo del exponencial de la entropía de Shannon y q=2 cuantifica los taxones dominantes y corresponde el inverso del índice de concentración de Simpson (Chao et al. 2014). La representatividad de los muestreos se exploró mediante las curvas de rarefacción de la abundancia de los taxones. Estos análisis de diversidad se realizaron en el programa R, versión 4.1.1 (R Core Team, 2020) empleando el paquete de iNEXT. Adicionalmente, se realizó un análisis de componentes principales (ACP) en el cual se asociaron los porcentajes de los ítems alimenticios y los taxones, el análisis se realizó en el programa R, versión 4.1.1 (R Core Team, 2020) empleando el paquete Factoextra.
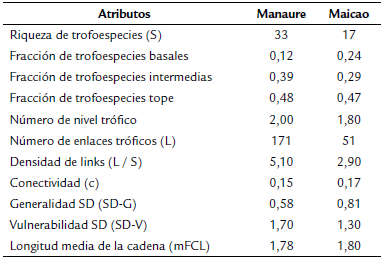
Para calcular las propiedades de las redes tróficas, se construyeron matrices binarias depredador - presa, usando los datos obtenidos según el análisis de contenido estomacal y revisión de literatura de la dieta de los taxones que no se diseccionaron (Muñoz, 2009), lo cual se hace para no excluir a los organismos de los análisis, debido a que esto produciría variación en los cálculos de las propiedades de las redes tróficas. A cada red trófica se le estimaron las siguientes propiedades: riqueza de especies tróficas o trofoespecies (S, que es el número de trofoespecies), número de nivel trófico (posición trófica ocupada por las trofoespecies), número de enlaces tróficos (L, cada una de las interacciones de depredador-presa y consumidor-recurso a través de la identificación del contenido estomacal), densidad de links (L / S, número promedio de enlaces de alimentación por trofoespecie) y la conectividad (c, proporción de posibles enlaces que se dan entre las trofoespecies) (Dunne et al. 2002). De igual manera se estimó la generalidad SD (SD-G, número promedio de presas por consumidor), la vulnerabilidad SD (SD-V, número promedio de consumidores por presa), la longitud media de la cadena (mFCL), número de taxones basales (b, taxones que solo son presas), número de taxones intermedios (i, taxones que son presas y consumidores) y número de taxones superiores (t, taxones que solo son consumidores). Los modelos de las redes tróficas se realizaron en el programa Network3D (Williams, 2010).
RESULTADOS
Caracterización fisicoquímica de las charcas
La menor temperatura del agua se observó en Maicao (31,9 ± 0,14 C°) y la mayor en Manaure (32,2 ± 0,7 C°). El pH en las dos charcas presentó valores ligeramente básicos (8,15 y 7,97 en Manaure y Maicao respectivamente). El mayor registro de oxígeno disuelto estuvo en Manaure (6,3 ± 2,9 mg/L). La charca de Maicao tuvo los mayores valores de nitritos (0,89 ± 0,06 µg/L) y nitratos (0,28 ± 0,01 µg/L) en la charca de Manaure mostró los mayores valores de fosfatos (0,51 ± 0,01 mg/L) (Tabla 1).
Estructura de comunidades
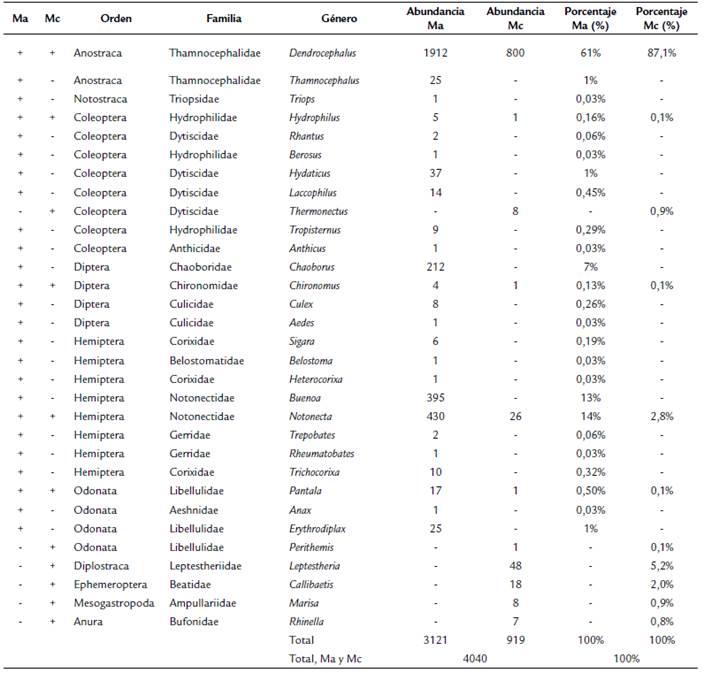
Se recolectaron 4040 individuos, representados en 10 órdenes, 17 familias y 31 géneros (Tabla 3). En Manaure se analizaron 123 estómagos pertenecientes a 25 géneros (°D). Los más abundantes fueron: Dendrocephalus (Anostraca) con 1912 individuos (61 % de la abundancia total: la mayor abundancia), Notonecta (Hemiptera) con 430 individuos (14 %), Buenoa (Hemiptera) con 395 (13 %) y Chaoborus (Diptera) con 212 (7 %). En Maicao se analizaron 62 estómagos pertenecientes a 11 géneros ( 0 D), los más abundantes fueron: Dendrocephalus (Anostraca) con 800 individuos (87 % de la abundancia total: la mayor abundancia), Leptestheria (Diplostraca) con 48 (5 %) y Notonecta (Hemiptera) con 26 individuos (3 %).
Tabla 3 Lista de géneros identificados en las charcas de Manaure y Maicao con sus abundancias y porcentajes de abundancia. Ma: Manaure, Mc: Maicao. + indica la presencia del taxón; - indica la ausencia del taxón
En Manaure se determinaron cuatro géneros con similares abundancias (D): Dendrocephalus, Notonecta, Buenoa y Chaoborus; y en Maicao se registraron 2 géneros con abundancias similares (1D): Dendrocephalus y Leptestheria; y 1 género dominante (2D): Dendrocephalus. La cobertura de muestreo en Manaure y Maicao fueron de 99,7 % y 99,6 % respectivamente, lo cual indica que fueron representativos; no obstante, los intervalos de confianza de la riqueza en ambos sitios fueron amplios (Fig. 1).
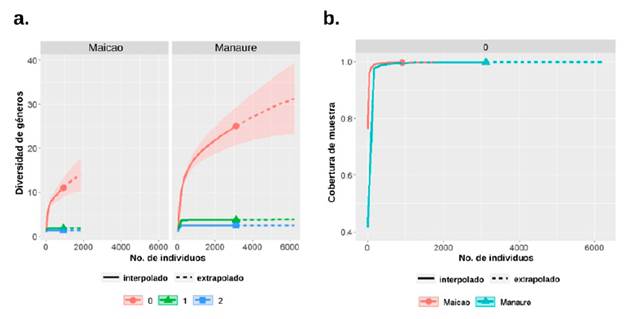
Figura 1 Perfiles de diversidad (qD) y cobertura de muestreo en función a la abundancia. (a) Curva de rarefacción basada en la riqueza de taxones, los números indican el orden (q) de la diversidad; 0 = riqueza, 1 = géneros igualmente abundantes, 2 = géneros dominantes. (b) Cobertura de los muestreos en función de la abundancia en las charcas de Maicao y Manaure.
En Manaure, el análisis de componentes principales (ACP) acumuló el 85,8 % de la variabilidad de la información en los dos primeros ejes (Fig. 2a). Se evidenció asociación entre los odonatos Pantala y Erythrodiplax y el coleóptero Hydaticus con altas proporciones de material animal (MA: 16,8 % ± 4,3 %; Suplemento 3) que son considerados como taxones tope de la red trófica. Laccophilus (Coleoptera) se asoció con un alto consumo de microalgas (6,6 %) en comparación con los demás géneros; Culex (Diptera) exhibió el mayor consumo de MA sin identificar (34,6 %), los anostracos Dendrocephalus y Thamnocephalus tuvieron altas proporciones de materia orgánica particulada fina (MOPF: 91,6 % ± 0,1 %); mientras que Hydrophilus presentó altas proporciones de materia orgánica particulada gruesa (MOPG: 43,2 % ± 9,50 %), el cual a su vez presentó el mayor porcentaje de consumo de este recurso junto con Chironomus (Diptera) (Fig 2a). Por su parte, Tropisternus (Coleoptera) y Chaoborus (Diptera) no mostraron asociaciones con ninguno de los recursos en particular.
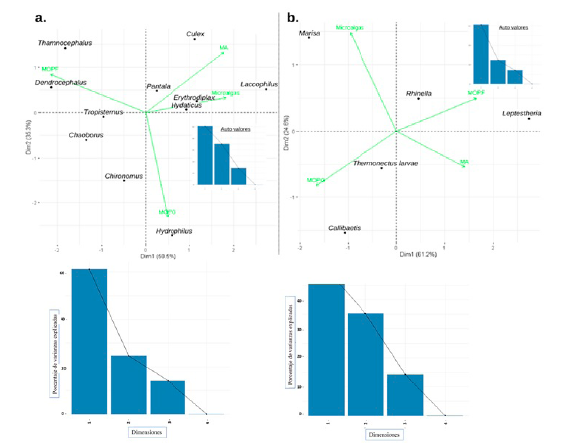
Figura 2 Análisis de componentes principales (ACP) y auto valores en las charcas de Manaure (a) y Maicao (b) a partir de los porcentajes de análisis de contenido estomacal. Material animal (MA), materia orgánica particulada gruesa (MOPG) y materia orgánica particulada fina (MOPF).
En Maicao el ACP acumuló el 85,8 % de la variabilidad de los datos en los dos primeros ejes (Fig. 2b). En esta charca, Marisa (Mesogastropoda) presentó una asociación directa con el consumo de microalgas (8,8 %), mientras que los renacuajos de Rhinella (Anura) y Leptestheria (Diplostraca) se asociaron con la MOPF (90,5 % ± 1,9 %); por su parte, Callibaetis (Ephemeroptera) y larvas de Thermonectus (Coleoptera) lo hicieron con la MOPG (12 % ± 2,1 %); no obstante, Callibeatis y Lepthesteria presentaron considerables porcentajes de MA en sus estómagos (2,4 % y 4,2 % respectivamente), con relación a los demás ítems (Fig. 2b).
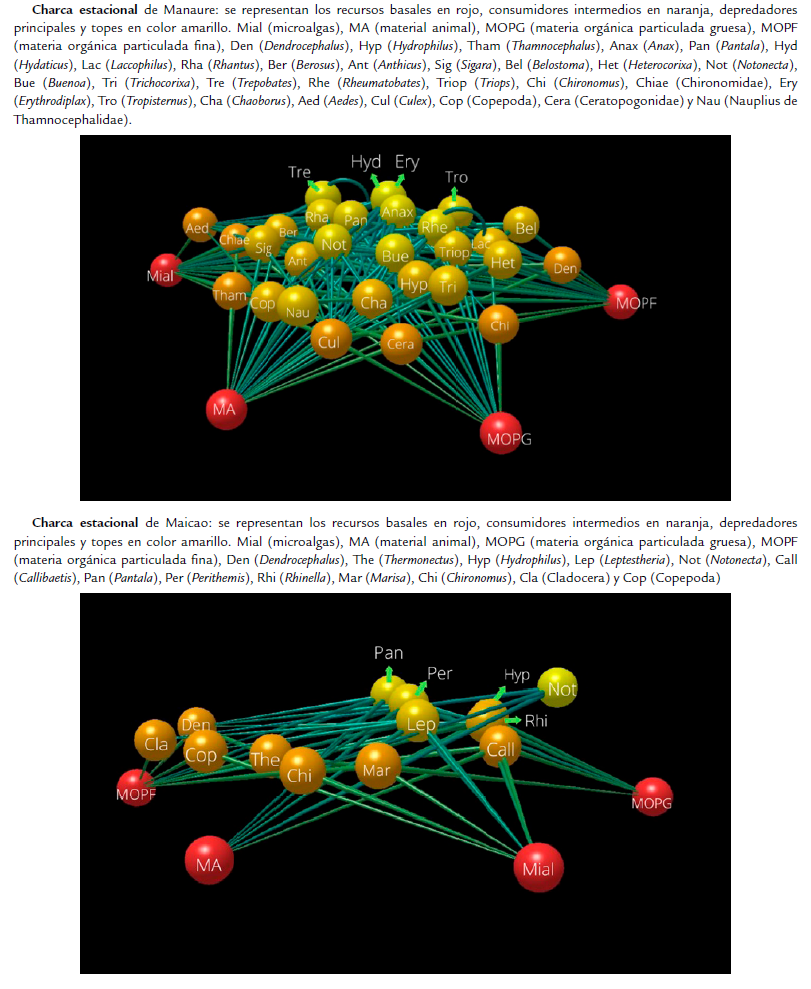
Con base en los hábitos alimenticios se establecieron cuatro categorías tróficas: carnívoros, herbívoros, omnívoros y detritívoros, y las fuentes basales, a las cuales se les llamó productores. A partir de estas categorizaciones obtenidas de los contenidos estomacales y la revisión de literatura, se presentaron dos niveles tróficos en las redes por encima de los productores y recursos; consumidores de primer orden (detritívoros, herbívoros, omnívoros y carnívoros) y consumidores de segundo orden (omnívoros y carnívoros) (Fig. 3).
En ambas redes tróficas de las charcas se observaron los mismos recursos basales [microalgas (Mial), materia orgánica particulada fina (MOPF), materia orgánica particulada gruesa (MOPG) y material animal (MA)], así como también se registraron detritívoros como consumidores de primer orden como Dendrocephalus y Thamnocephalus en Manaure (Fig. 3a) y Chaoborus y Callibaetis en Maicao (Fig. 3b). Herbívoros como Marisa y omnívoros como Anthicus y Chironomus. También se registraron consumidores carnívoros de segundo orden como Pantala, Erythrodiplax, Anax, Tropisternus, Rhantus, Notonecta, Buenoa, Trepobates, Rheumatobates en Manaure (Fig. 3a) y Perithemis, Hydaticus, Pantala y Notonecta en Maicao (Fig. 3b); hasta organismos omnívoros (mayormente detritívoros) de segundo orden como lo es el caso de Lepthestheria en la charca de Maicao (Fig. 3b). De igual manera, dentro de los consumidores de segundo orden se identificaron depredadores tope para las dos charcas, los cuales fueron: Hydaticus y Tropisternus en los coleópteros, Pantala, Perithemis, Erythrodiplax, Anax en los odonatos y Notonecta y Trepobates en los hemípteros (Fig. 3 a,b ).
La riqueza de trofoespecies en Manaure (33) casi que duplica a la de Maicao (17); así como el número de enlaces tróficos (171) y densidad de links (5,10). No obstante, el número de niveles tróficos (Manaure: 2,00; Maicao: 1,80), la longitud media de la cadena (Manaure: 1,78: Maicao: 1,80) y la conectividad (Manaure: 0,15; Maicao: 0,17) fueron similares. Maicao presentó el doble de la fracción de trofoespecies basales (0,24), pero una menor fracción de las intermedias (0,29); no obstante, las dos charcas tuvieron una fracción similar de trofoespecies tope (Manaure: 0,48; Maicao: 0,47). La vulnerabilidad en Manaure fue mayor (1,70) que en Maicao (1,30). (Tabla 2).
DISCUSIÓN
La charca de Manaure presentó una red trófica más compleja que la de Maicao, por los valores de las propiedades de las redes. El mayor número de enlaces tróficos, densidad de links y el canibalismo por parte de Tropobates y Tropisternus observado en Manaure, fueron las principales propiedades que diferenciaron las redes; no obstante, la longitud de la cadena fue similar en los dos ecosistemas, lo cual podría variar si se tienen en cuenta peces anuales que ya han sido registrados en ecosistemas temporales cercanos (Tamaris-Turizo et al. 2020) y siendo más detallados en la identificación de los tejidos de restos de animales dentro de estómagos que no se lograron identificar; de igual manera sucedió con la conectividad y el número de taxones superiores.
Algo que pudo influir en el menor número de enlaces en Maicao es la intervención e impacto humano en dicha charca, debido a que a diferencia de Manaure se encontraba mucho más cerca de la carretera, y en su momento más expuesta a residuos y contaminantes. Tamaris-Turizo y colaboradores en 2018 mediante la evaluación de redes tróficas en un gradiente altitudinal a lo largo de un río de la Sierra Nevada de Santa Marta observaron que los impactos humanos se asociaron con redes con un menor número de enlaces y por ende menor complejidad.
Esta misma tendencia se evidenció con el análisis de diversidad. Los datos de riqueza fueron similares a los registrados por Serna-Macías et al. (2019) con 28 géneros y Tamaris-Turizo et al. (2020) con 30 géneros, pero difieren de Maicao. Esto pudo ocurrir porque las composiciones de las charcas estacionales tienden a ser cambiantes e inestables por la dependencia de las precipitaciones (Williams, 2006). En la localidad de Maicao se identificaron algunos géneros que no estuvieron presentes en Manaure, como fue el caso de Marisa (Mesogastropoda, Ampulariidae), el cual es una especie exótica e invasora gran consumidora de la vegetación acuática (Catálogo Español de EEI, 2013), la cual estuvo más asociada a plantas de la familia Araceae como Lemna minor L y Onagraceae como Ludwigia helminthorrhiza; sin embargo, es recomendable análisis que asocien dichas relaciones planta-animal y el efecto del impacto antrópico mediante la introducción de especies invasoras.
Los insectos y crustáceos son considerados como la fauna dominante en este tipo de charcas estacionales (Williams, 2006) de manera general y entre los insectos los órdenes más representativos de la biota hídrica temporal son Hemiptera, Coleoptera, Trichoptera y Chironomidae (Diptera), lo cual coincide parcialmente con lo observado en este tipo de sistemas en España (Sancho y Lacomba 2010) y en otras charcas estacionales del departamento de La Guajira (Serna-Macías et al. 2019; Tamaris-Turizo et al. 2020), exceptuando la presencia de Trichoptera, orden no registrado hasta la fecha en estos ecosistemas del norte de Colombia. Los insectos dominantes en las charcas evaluadas fueron Hemiptera, Diptera y Coleoptera, entre los dípteros dominó Chaoborus (Chaoboridae), y en los crustáceos Dendrocephalus, el cual fue el taxón con mayores abundancias para las dos charcas. Este último género se registró previamente en estudios para el departamento de la Guajira, Colombia (Cohen et al. 2014; Serna-Macías et al. 2019; Tamaris-Turizo et al. 2020) así como en Venezuela (García, 1997), Argentina (Pereira y Ruiz, 1995) y Brasil (Rogers et al. 2012).
Las charcas evaluadas se encuentran cerca de zonas agrícolas y ganaderas al igual que a las carreteras, esto puede influenciar a los valores fisicoquímicos obtenidos. Serna-Macías y colaboradores en 2019 mencionan que los altos valores de nitratos y nitritos registrados en su estudio, pueden relacionarse con la fertilización de estas zonas agrícolas y sus arrastres a causa de las lluvias. En nuestro estudio, los valores de nutrientes fueron similares a los registrados en la charca de Maicao, pero totalmente distintos con respecto a la de Manaure. Los valores de amonio fueron menores en las dos charcas con respecto al estudio mencionado, mientras que los registros de pH fueron un poco más altos por encima de la neutralidad. Cabe destacar que en el estudio de Serna-Macías et al (2019), en los periodos de bajas precipitaciones las concentraciones de amonio aumentaron, lo cual puede estar relacionado con la mayor acumulación de materia orgánica en estos periodos, pero, para corroborar esto habría que hacer correlaciones teniendo en cuenta las precipitaciones y los valores de los nutrientes.
Los análisis de las dietas mostraron una baja abundancia de omnívoros, algunos representantes fueron: Triops, Chironomus y Anthicus en Manaure y Rhinella y Leptestheria en Maicao. No obstante, estos resultados difieren de lo registrado por Muñoz et al. (2009) y Williams (2006). Esta discrepancia podría deberse a que estas charcas se secan durante algunos meses durante el año, por ende, los organismos deben aprovechar los recursos alimenticios disponibles. Adicionalmente, la corta longitud de la cadena deja en evidencia los pocos niveles tróficos presentes, lo cual está asociado con el alto grado de especialización dietario de los organismos.
En las dos redes tróficas, la MOPF fue el recurso más abundante, excepto en hemípteros heterópteros por su hábito alimenticio (Vivas y López, 2013). De igual manera, indica que es posible que el alto consumo de la MOPF en taxones carnívoros sensu stricto sea porque dicho material era el recurso principal de alimentación de su presa, además de que una gran cantidad de la MOPF proviene del fraccionamiento de la MOPG (Allan y Castillo, 2007).
Andramunio y Caraballo (2012) mencionan que los sistemas acuáticos inundables reciben un gran aporte de materia orgánica de fuentes externas que sirve como fuente de alimento a organismos que consumen detritos, gracias a que toda esta materia orgánica experimenta una degradación microbiana y como consecuencia presenta una alta disponibilidad del recurso (Tomanova et al. 2006).
Los principales depredadores en las dos charcas fueron Coleoptera, Odonata y Hemiptera, lo cual coincide con estudios realizados en charcas estacionales del departamento de la Guajira, Colombia (Serna-Macías et al. 2019), en la localidad de Tenería (Barquisimeto, Venezuela) (García, 1997), en la Comunidad Valenciana, España (Sancho y Lacomba 2010) y coincide parcialmente con las abundancias registradas en un estudio de Uruguay (Illarze et al. 2021) en donde los Coleópteros fueron el grupo más abundante de las charcas estudiadas.
El recurso que presentó una mayor vulnerabilidad (SD-V) fue la MOPF, debido a que fue el ítem más consumido por la mayoría de macroinvertebrados. Esta vulnerabilidad es común en las redes tróficas de los sistemas acuáticos, porque su energía depende de las fuentes de materia orgánica (Amadeu-Santana et al. 2015) como la gruesa (Tamaris-Turizo y Rodríguez-Barrios 2015), lo que confirma que la vía de los detritos es de mayor importancia en estos sistemas al compararla con la de los productores.
CONCLUSIONES
Los resultados de esta investigación y de investigaciones anteriores en la Guajira sugieren que en estos ecosistemas temporales se encuentra una alta riqueza de macroinvertebrados. Las redes tróficas desarrolladas a partir de los análisis de dieta en los organismos resaltan la importancia de la MOPF como un recurso alimenticio dominante en los macroinvertebrados y vertebrados acuáticos. Del mismo modo, a partir de los resultados obtenidos con esta investigación se observan estructuras tróficas disímiles entre los dos sistemas evaluados que sirven como grandes aportes para el conocimiento inicial de las relaciones alimenticias de charcas estacionales en el departamento de la Guajira. Sin embargo, es necesario seguir llevando a cabo estudios que nos permitan conocer más la biodiversidad que habita en estos ecosistemas temporales en el departamento de la Guajira, así como características más específicas de las estructuras tróficas a través de redes tróficas con más detalle taxonómico posible (género y especie) y análisis de isotopos estables, para que permiten rastrear las fuentes de energía y estimar los porcentajes de asimilación de estas. Por último, se recomienda realizar comparaciones de redes tróficas de más charcas estacionales disponibles a lo largo del departamento de la Guajira y tener en cuenta las diferentes épocas climáticas, así como el proceso de llenado y vaciado de las charcas, ya que las precipitaciones activan procesos del desarrollo en algunos organismos (ej. la eclosión), lo que podría generar variación en la composición biológica y consecuentemente en las propiedades de las redes tróficas. A los evaluadores anónimos por sus comentarios para la mejora del manuscrito.