










Services on Demand
Journal
Article
 Spanish (pdf)
Spanish (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mailIndicators
-
 Cited by SciELO
Cited by SciELO -
 Access statistics
Access statistics
Related links
-
 Cited by Google
Cited by Google -
 Similars in
SciELO
Similars in
SciELO -
 Similars in Google
Similars in Google
Share

 Permalink
PermalinkRevista Salud Uninorte
Print version ISSN 0120-5552
Salud, Barranquilla vol.31 no.1 Barranquilla Jan./Apr. 2015
Efecto de las isoflavonas de la soja en la salud ósea de adultos y niños
The effects of soybean isoflavones over the bone health of adult and children
Dary Luz Mendoza Meza1, Carlos Roa Mercado2, César Ahumada Barraza3
1 Química Farmacéutica. MSc. Bioquímica. Docente del Programa de Especialización en Química Orgánica, Facultad de Ciencias Básicas, Universidad del Atlántico. Barranquilla (Colombia).
2 Licenciado en Biología y Química. Especialista en Química Orgánica, Universidad del Atlántico. Barranquilla (Colombia).
3 Licenciado en Biología y Química. Especialista en Química Orgánica, Universidad del Atlántico. Barranquilla (Colombia).
Correspondencia: Dary Luz Mendoza Meza. Grupo de Investigación en Productos Naturales y Bioquímica de Macromoléculas, Universidad del Atlántico. Km 7, vía a Puerto Colombia (Barranquilla-Atlántico). darymendoza@mail.uniatlantico.edu.co
Fecha de recepción: 30 de septiembre de 2014
Fecha de aceptación: 5 de noviembre de 2014
Resumen
Las isoflavonas de la soja son metabolitos secundarios conocidos como fitoestrógenos por su similitud estructural con el 17b-estradiol. La daidzeina y la genisteina son las principales isoflavonas de la soja. Debido a su capacidad de unión a los receptores de estrógeno (RE) >son consideradas moduladores selectivos de los receptores de estrógenos (MSRE). El consumo dietario de isoflavonas es relacionado con la disminución de los síntomas de la menopausia, prevención de enfermedades cardiovasculares y de otras enfermedades crónicas como la diabetes mellitus. Las isoflavonas han sido propuestas también para el tratamiento de algunos tipos de cáncer y la prevención de la osteoporosis; sin embargo, existe gran controversia sobre los posibles efectos estrogénicos en niños púberes y prepúberes alimentados con fórmulas a base de proteína de soja. El objetivo de este trabajo es presentar una revisión sistemática de la química de las isoflavonas de la soja y sus efectos sobre la salud del hueso en adultos y niños.
Palabras clave: isoflavonas de la soja, genisteína, daidzeína, salud ósea.
Abstract
The soy isoflavones are secondary metabolites known as phytoestrogens because of its structural similarity to 17b-estradiol. Genistein and Daidzein are the main soy isoflavones. Due their binding ability to estrogen receptors (ER) they are considered as selective estrogen receptor modulators (SERM). The dietary intake of isoflavones has been associated with decreasing menopause symptoms, prevention of cardiovascular disease and some chronic diseases such as diabetes mellitus. Also, the isoflavones have been proposed for the treatment of certain kind of cancer and osteoporosis prevention, but there is a lot of controversy about the possible estrogenic effects in pubertal and prepubertal children who have been fed with soy protein-based formulas. The aim of this research is to present a systematic review of the soy isoflavone chemistry and their effects on bone health in adults and children.
Keywords: Soy isoflavones, genistein, daidzein, bone health.
Introducción
Algunos vegetales contienen compuestos fito-químicos no nutricionales que ejercen acciones protectoras sobre la salud humana. Un grupo importante de metabolitos secundarios son los fitoestrógenos o estrógenos vegetales, los cuales se clasifican en isoflavonas, lignanos y coumestanos (1).
La soja [Glycine max (L.) Merr] produce diversos tipos de isoflavonas y también produce coumestanos, cuando la planta está en crecimiento (2).
A las isoflavonas de la soja se les atribuye amplias propiedades terapéuticas, las cuales se han relacionado con la capacidad de estas moléculas para unirse a los receptores de es-trógenos (3, 4). Entre las propiedades de las isoflavonas se destaca su efecto en el metabolismo óseo, siendo relacionado con el mantenimiento de la densidad mineral ósea (DMO) en mujeres menopáusicas y posmenopáusicas (5). A pesar de la existencia de numerosos estudios que respaldan esta hipótesis existen otros que la descartan (6), siendo este un tema muy controversial. Entre las publicaciones más recientes se encuenta un estudio doble ciego al azar realizado con mujeres taiwanesas, tratadas durante dos años con 300 mg/día de isoflavonas, en el cual no se observó diferencia en DMO de la espina lumbar o el fémur entre el grupo tratamiento y el grupo placebo (7); resultados similares se observaron en dos estudios independiente realizados recientemente con mujeres americanas postmenopáusicas (8) y menopáusicas (9).
Además del efecto sobre la DMO, las isoflavonas de la soja también han sido reportadas por su posible acción anticancerígena en tejidos de mama (10), próstata (11) y colón (12); también como mitigante de los efectos adversos de la radioterapia sobre tejidos normales (13), protector del endotelio vascular (14), promotor de la proliferación y la diferenciación de células de la dentina (15), y coadyuvante en el tratamiento de la diabetes mellitus (16) y de enfermedades renales (17). Actualmente se está estudiando su efecto preventivo en el Alzheimer (18, 19).
A pesar de las propiedades beneficiosas atribuidas a las isoflavonas de la soja, existe gran debate sobre los posibles efectos estrogénicos en niños púberes y prepúberes, y especialmente sobre los efectos en el crecimiento óseo.
El propósito de este trabajo es presentar una revisión sistemática de la química de las isoflavonas de la soja y los resultados de estudios científicos sobre los efectos de las isoflavonas en la salud ósea de los humanos.
Química de las isoflavonas de la soja
Las isoflavonas se caracterizan por tener peso molecular y estructura química parecida al 17b-estradiol, una hormona esteroide femenina encargada de regular funciones sexuales, reproductivas y del metabolismo óseo. Las isoflavonas derivan biosintética-mente de la unión de un precursor aromático (hidroxicinamil coenzima-A éster) con otro alifático (malonil coenzima-A). La característica estructural básica de las isoflavonas es el núcleo flavona, compuesto por dos anillos bencénicos (Ay B) unidos por un anillo pirano heterocíclico (C) (20, 21) (figura 1).
Una de las propiedades químicas más importantes de las isoflavonas es su gran poder antioxidante, atribuido a la capacidad del grupo fenólico de captar y remover los radicales libres (22, 23).
Existen por lo menos 220 especies vegetales que contienen isoflavonas, siendo las leguminosas la de mayor contenido (24). La soja es la fuente más importante de isoflavonas en la dieta humana (25); contiene 12 isoflavonas diferentes clasificadas en dos componentes, b-glucósidos y agliconas. Las isoflavonas b-glucósidos se caracterizan por tener una cadena de glucosa en su estructura, mientras que la aglicona no tiene glucosa.

Las isoflavonas de la soja más investigadas desde el punto de vista biológico y farmacológico son la genisteína (nombre IUPAC: [5,7- dihidroxi-3-(4-hidroxifenil)-4H-1-benzopiran-4-ona] y la daidzeína (nombre IUPAC: [7-hidroxi-3-(4-hidroxifenil)-4H-1-benzopiran-4-ona] (22).
La genisteína tiene peso molecular de 270.24 Dalton y punto de fusión entre 200-202 °C, es un sólido cristalino de color amarillo, insoluble en agua y bastante soluble en etanol.
La daidzeína tiene peso molecular de 254.24 Dalton y punto de fusión entre 96-98 °C, también se presenta en estado sólido y es insoluble en agua.
Tanto la genisteína como la daidzeína se encuentran de forma natural como 7-b-glucósidos, y reciben el nombre de genistina y daidzina, respectivamente. La unión de estas isoflavonas a un azúcar aumenta su solubilidad en agua y las hace más estables a las condiciones fisiológicas normales (26).
En la soja el contenido de daidzeína y la genisteína es alto; se ha reportado alrededor de 0.1 a 0.3 mg/g de soja (27), aunque estas concentraciones pueden variar dependiendo de la ubicación geográfica, clima y condiciones de crecimiento de los cultivos.
En los alimentos a base de soja, la relación porcentual aproximada del b-glucósido/ aglicona con respecto al contenido de isoflavonas totales es: genistina/genisteína 50-55% y daidzina/ daidzeína 40-45% (28).
La figura 2 muestra la estructura general de una isoflavona aglicona y su respectivo glucósido (26).

Bioquímica de las isoflavonas la soja
En los humanos, las agliconas de isoflavona se absorben fácilmente en la parte superior del intestino delgado por difusión pasiva, alcanzando los niveles plasmáticos máximos después de una hora de ser ingerido. En contraste, las isoflavonas p-glucósidos no son absorbidas pasivamente a través del tracto gastrointestinal; para que la absorción ocurra, estas deben ser hidrolizadas hasta sus agliconas, las cuales son más hidrofóbicas y tienen peso molecular más bajo.
La hidrólisis de las isoflavonas p-glucósidos es catalizada por enzimas p-glucósidasas de la mucosa intestinal o por p-glucósidasas producidas por la flora intestinal bacteriana (29).
En individuos con deficiencia en la enzima lactasa-florizina hidrolasa (enzima responsable de la hidrólisis de la lactosa), la digestión de las isoflavonas p -glucósidos disminuye en el intestino delgado (30).
Las isolavonas p-glucósidos no hidrolizadas en el intestino delgado pasan al intestino grueso, donde las poblaciones de microorganismos son más altas; en este entorno más anaeróbico no solo se produce la hidrólisis, también ocurren modificaciones reductivas en el anillo heterocíclico de la isoflavona daidzeína, produciéndose dihidrodaidzeína y O-desmetil-angolensina (figura 3).
Se ha establecido que aproximadamente 30 % de la población posee una flora intestinal que es capaz de reducir la daidzeína a la isoflavona 7,4>-dihidroxi-isoflavona (también denominada equol); esta molécula tiene un centro quiral en el carbono 3 (C-3), razón por la cual se generan dos isómeros: el S-(-) equol y R-(-) equol; esto es importante porque el isómero R del equol es 30 veces más débil en su afinidad por el receptor de estrógeno beta (RE-b) comparado con el isómero S (31, 32).

La importancia bioquímica de la producción de S-(-) equol se ha evidenciado en estudios realizado en mujeres japonesas postmeno-páusicas no productoras de este metabolito, a quienes se les administró una dosis de 10 mg/día de S-(-) equol por un año, y se observó una disminución de la excresión urinaria de deoxypyridinoline, un marcador de la reabsorción ósea (33).
Otro derivado de la daidzeína es la puerarina (daidzeína-8-C-glucósido), la cual es resistente a la hidrólisis enzimática, debido al enlace de C-C entre la glucosa y la isoflavona; como consecuencia de ello, la puerarina es llevada a través de transportadores de glucósidos al interior de la célula intestinal, donde se convierte en p-glucurónidos, reacción que es catalizada por las UDP-glucuronil transferasas. También se convierte en ésteres de sulfato, reacción catalizada por PAPS (3'-fosfoadenosina-5'-fosfosulfato)- sulfotransferasas (34) (figura 4).
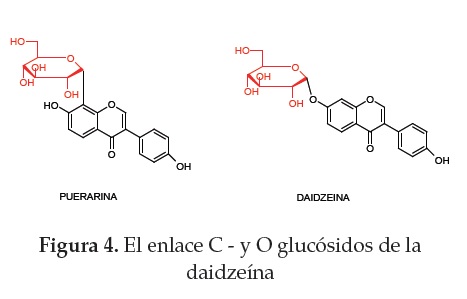
Una vez absorbidas, las isoflavonas agliconas se distribuyen más fácilmente a los tejidos periféricos, comparadas con las isoflavonas b-glucosidos (35). Las isoflavonas glucorinadas y sulfatadas entran a la circulación sistèmica a través de la circulación entero hepática (36).
El metabolismo de las isoflavonas también puede ocurrir localmente, en la vecindad de las células inflamatorias (macrófagos, neutrófilos y eosinófilos activados), donde cumplen una función importante al reaccionar con especies oxidantes reactivas que se generan bajo condiciones fisiológicas, como el ácido hipocloroso (HOCl) y el peroxinitrito (ONO2). Los derivados clorinados y mononitrados son posteriormente conjugados con el ácido glucorónico y excretados en la bilis (36, 37). Estas reacciones explicarían las propiedades antioxidantes y antiinflamatorias de las iso-flavonas in vivo (38, 39).
En los tejidos, la genisteína, la daidzeína y algunos de sus derivados metabólicos pueden unirse a los receptores de estrógenos, razón por la cual se les ha catalogado como moduladores selectivos de receptores de estrógenos (MSRE). En algunas células la unión de los MSRE a los receptores de estrógenos simula la actividad del 17b-estradiol (E2), pero esto no es general, ya que en otras células el efecto es contrario (40).
Para entender cómo las isoflavonas de la soja pueden influir en el crecimiento óseo es necesario revisar los efectos del estrógeno en el hueso.
Estrógenos y el crecimiento óseo
El volumen del hueso se mantiene gracias a dos procesos de remodelación: uno es la reabsorción ósea, llevada a cabo por los os-teoclastos, y el otro es la formación del hueso, llevada a cabo por los osteoblastos (41, 42).
La remodelación ósea está controlada por diversas hormonas, como la paratohormona, también por citoquinas como la interleucina-1 (IL-1), interleucina-6 (IL-6), por glucocorticoides y hormonas esteroides.
Un desequilibrio entre la actividad osteoblás-tica y osteoclástica conduce a enfermedades metabólicas en los huesos; por ejemplo, la disminución de estrógenos después de la menopausia está asociada con la pérdida ósea y la osteoporosis (43, 44).
Debido a su carácter hidrofóbico, los estró-genos pueden difundir dentro y fuera de las células, sin embargo, en las células dianas los estrógenos son retenidos con gran afinidad y especificidad por unas proteínas de unión intracelulares llamadas receptores de estró-geno (RE). Cuando los estrógenos se unen a los RE forman un dímero, que induce a una respuesta biológica interactuando con la cromatina y modulando la trascripción de los genes (45, 46).
A la fecha se han caracterizado dos tipos de RE: el receptor alfa (RE-a), presente en osteo-blastos y osteoclastos y, el receptor beta (RE-b), expresado solo por osteoblastos (figura 5).

Aunque las bases moleculares del efecto os-teoprotector del estrógeno son desconocidas, se han planteado varias hipótesis que tratan de explicar el papel de esta hormona en la remodelación del hueso en algunas especies animales.
La detección de los RE-a en los osteoblastos y osteoclastos sugirió que las acciones directas del estrógeno en ambos tipos de células son responsables de los efectos beneficiosos sobre el esqueleto.
Experimentos publicados por Nakamura y colaboradores en 2007 muestran que el estrógeno previene la pérdida del hueso a través de la inhibición del desarrollo de los osteoclastos; en este trabajo el estrógeno indujo la apoptosis y sobrerregulación de la expresión de señales de muerte celular mediadas por el ligando Fas (FasL) en os-teoclastos de huesos trabeculares de ratones silvestres que expresan el RE-a, pero no en ratones con el RE-a truncado (RE-aDeltaOc/ DeltaOc); resultado que soporta un modelo en el cual el estrógeno regula la vida media de los osteoclastos maduros vía inducción de la señal de muerte Fas/FasL (47). Esta hipótesis es respaldada por los estudios de García y colaboradores en 2013, quienes observaron que el E2 induce la apoptosis de las células osteoclásticas a través de la escisión y la solu-bilización del ligando Fas (FasL), expresado por los osteoblastos (48). La supresión de la resorción ósea osteoclástica conduce a la disminución de la remodelación ósea y, por consiguiente, el aumento de la masa ósea y la fuerza, proporcionando de esta forma una explicación para la función osteoprotectora de los estrógenos y de los MSRE.
Aunque existen evidencias de la participación de los estrógenos en la diferenciación celular de los osteoblastos (49, 50), la función del RE-a en este linaje celular ha sido más difícil de explicar.
Almeida y colaboradores en 2013 realizaron experimentos con células osteoblásticas de ratón en diferentes estados de diferenciación, y encontraron que el RE-a cumple una función esencial en la acumulación y mantenimiento de los huesos a través de acciones en células autónomas de progenitores de osteoblastos y osteoclastos. Estos autores proponen que el papel del RE-a en los osteoblastos y los osteoclastos es diferente en compartimentos óseos distintos: en los osteoblastos RE-a promueve la formación de hueso en la superficie periosteal y previene la reabsorción en la superficie endocortical, mientras que en los osteoclastos, el RE-a impide la reabsorción de hueso esponjoso (51).
En niños púberes se ha demostrado que el estrógeno tiene un efecto bifásico sobre el crecimiento epifisario del hueso, lo cual significa que los efectos cambian con el tiempo. Un estudio realizado por Cutter en 1997 mostró que dosis de estrógenos correspondientes a niveles de estradiol sérico de ~4 pg/mL (15 pmol/L) causan un aumento de más del 60 % con respecto a la tasa de crecimiento prepuberal en niños y niñas (52).
En los varones, informes de osteopenia severa se han asociado positivamente con mutaciones en el gen para el receptor de estrógeno (ER-a) y deficiencia en la aromatasa, la enzima que cataliza la conversión de los andrógenos en estrógenos; estos hallazgos plantearon el interrogante de cuál es la función de los estrógenos en el esqueleto del varón.
La administración de un inhibidor de la enzima aromatasa (la testolactona) a niños con el síndrome de pubertad masculina precoz familiar mostró el efecto de la testolactona en la normalización tanto de la velocidad de crecimiento y la maduración ósea, a pesar de que los niveles de testosterona en suero se mantuvieron dentro del rango del varón adulto (53); otros experimentos respaldaron la observación de que niveles bajos de estrógenos estimulan el crecimiento y la maduración de hueso, lo cual podría explicar la maduración epifisaria más rápida de las niñas prepúberes en comparación con los niños (54). Publicaciones más recientes muestran que los estrógenos tiene una influencia significativa en la DMO en los niños y niñas, lo cual sugiere que hay un periodo de edad crítico para la acumulación de la masa ósea (55). Adicionalmente, los datos presentados por Khosla y colaboradores en 2001 indican que el estrógeno juega un papel clave tanto en la adquisición del pico de masa ósea en hombres jóvenes como en la pérdida ósea en hombres de edad avanzada; también sugieren que las disminuciones relacionadas con la edad, en niveles de estradiol biodisponible menores de 40 pmol/L, pueden ser la causa principal de la pérdida de masa ósea en los hombres de edad avanzada (56).
El descubrimiento de que los estrógenos se unen a los dos subtipos de receptores, RE-a y RE-b, se ha traducido en importantes esfuerzos para desarrollar ligandos que sean agonistas selectivos de estas moléculas, los cuales podrían tener un gran potencial terapéutico en un número de síntomas y/o enfermedades asociadas con la deficiencia de estrógenos, incluyendo los sofocos, osteoporosis y problemas cardiovasculares; adicionalmente, la sugerencia de que el RE-a promueve la proliferación celular y que los RE-p es anti-proliferativo puede ser relevante en el tratamiento de cáncer (57, 58).
Las isoflavonas de la soja y el crecimiento óseo
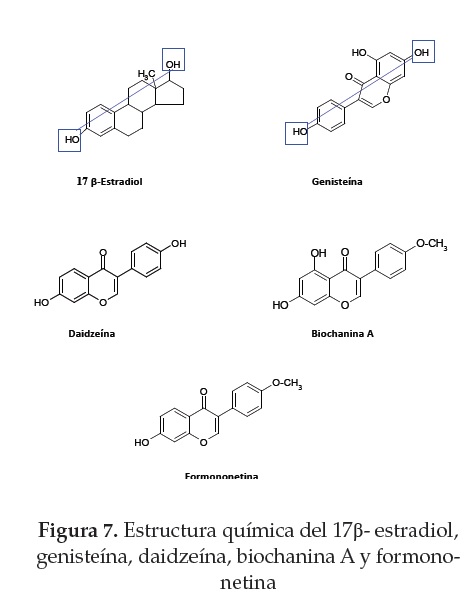
La estructura química de las isoflavonas es muy similar al E2 (20) (figura 8). El anillo aromático es un elemento clave para la unión con el RE. Si se compara la estructura del metabolito equol y el E2 se observa que son virtualmente superponibles, las distancias de los grupos hidroxilos son casi idénticas y determinan la interacción, mediante enlace de hidrógeno, con los aminoácidos del sitio de unión al receptor (3) (figura 6).

Aunque los mecanismos de unión del E2 y los fitoestrógenos son similares, la potencia transcripcional de estos últimos es más débil. Se ha sugerido que las exigencias estéricas del RE por el E2 (el estrógeno fisiológico primario) son la causa de la menor afinidad de las isoflavonas por el RE. La isoflavona de la soja, genisteína, se une tanto al RE-a como al RE-p, induciendo efectos estrogénicos con menor potencia que el E2 (~103-104 veces menor) (59). Al comparar la afinidad de la genisteína por los RE se observó que esta tiene una mayor afinidad por el RE-p, sin embargo, la pérdida de un grupo hidroxilo, como en el caso de la daidzeína y biocanina A, o dos hidroxilos, como en la isoflavona formononetina, disminuye la afinidad en la unión al receptor, lo cual indica que los grupos hidroxilos cumplen un papel fundamental en el reconocimiento ligando-receptor (figura 7).
La actividad estrogénica de las isoflavonas puede ser evaluada a través de la medición de la velocidad de unión del RE a los elementos de respuesta a estrógenos (ERE), lo que induce la activación de genes y la subsiguiente expresión de la actividad estrogénica.
Un estudio realizado con E2 y las isoflavonas genisteína, daidzeína, el equol y el coumes-trol demostró que la actividad estrogénica del E2 y las isoflavonas es dependiente de la concentración; adicionalmente, la hormona E2, así como el coumestrol y el equol, activan la unión de los RE-α y de los RE-β a los ERE, siendo el efecto ligeramente mayor a través del RE-ß comparado con el RE-a, por lo que la actividad estrogénica de la daidzeína es mediada a través de ambos isotipos de RE (RE-αy RE-β) en los individuos que son capaces de metabolizar la daidzeína a equol (60).
Debido a la capacidad de las isoflavonas de la soja de unirse a los RE, existe gran controversia sobre los posibles efectos estrogénicos de las isoflavonas en bebés y niños que son alimentados con fórmulas a base de soja, ya sea como suplemento o para reemplazar la leche humana o la leche de vaca.
Setchell y colaboradores publicaron un estudio en 1997 con niños de 4 meses de edad alimentados con fórmulas infantiles exclusivamente a base de soja (n= 7), leche de vaca (n= 7) o leche materna humana (n= 7), y encontraron concentraciones plasmáticas circulantes de isoflavonas en los infantes alimentados con fórmula a base de soja 13 000 a 22 000 veces mayor que las concentraciones plasmáticas de E2 en la vida temprana, lo cual puede ser suficiente para ejercer efectos biológicos, mientras que la contribución de las isoflavonas a partir de la leche materna y la leche de vaca fueron insignificantes (61).
Por otra parte, Giampietro y colaboradores (2004) mostraron que la alimentación a largo plazo con fórmulas a base de proteínas de soja, en la vida temprana no parece producir efectos hormonales similares al estrógeno (62).
Otros estudios han reportado la presencia de isoflavonas en orina, saliva y sangre en infantes alimentados con fórmulas a base de soja, sin embargo, no se describen consecuencias adversas (63).
Sobre el efecto de las isoflavonas de la soja en el metabolismo óseo del hombre, uno de los pocos estudios publicados reportó una posible relación entre el consumo de proteína de soja y el incremento en los niveles séricos del factor de crecimiento similar a la insulina (IGF-1), el cual está asociado con tasas mayores de formación de hueso. Este estudio fue realizado con hombres sanos (59.2 ± 17.6 años), quienes fueron alimentados con 40g/día de proteína de soja, durante 3 meses, en forma controlada y usando un diseño experimental a doble ciego, aleatorizado y paralelo; sin embargo, este mismo estudio no mostró incremento en los niveles séricos de otros marcadores de actividad osteoblástica, como son la actividad fosfatasa alcalina (ALP) y ALP específica del hueso (64).
El efecto de isoflavonas de la soja sobre el crecimiento óseo también ha sido evaluado en ratones y en cultivos in vitro de células osteoblásticas.
Estudios en el modelo animal ratón. El efecto de la genisteína en el metabolismo óseo de ratones hembras ovariectomizadas (OVX) ha sido comparado con el efecto del E2.
Un experimento realizado por Ishimi y colaboradores en 1999 mostró que ratones OVX a los cuales se les administró genisteína durante 2-4 semanas, en dosis de 0.1 a 0.7 mg/ día, restauraron completamente el volumen de hueso trabecular de la metáfisis distal del fémur, sin exhibir acción estrogénica en el útero, lo cual sugirió la utilidad de los fitoestrógenos para prevenir la pérdida ósea causada por la deficiencia de estrógenos (65).
Estudios posteriores mostraron que el efecto protector del hueso es dependiente de la dosis de genisteína y que dosis altas podrían inducir a hipertrofia uterina (66).
El efecto de las isoflavonas de la soja sobre la DMO también ha sido observado en ratones machos.
Fujioka y colaboradores en 2007 publicaron los resultados de su estudio con ratones machos y hembras sexualmente inmaduros, los cuales fueron alimentados con dietas suplementadas con isoflavonas de soja, y observaron que la daidzeína tiene un efecto específico, sexualmente dismórfico en la formación del hueso y la DMO durante el periodo de crecimiento de estos ratones; además los análisis histológicos revelaron que la velocidad de formación del hueso fue significativamente más alta en los machos alimentados con dietas suplementadas con daidzeína y genisteína comparado con el grupo control alimentados con dietas sin isoflavonas; en las hembras se observó que la velocidad de crecimiento fue más baja comparada con el grupo control (67).
Otro estudio realizado recientemente con ratones machos sugirió que dietas suple-mentadas con proteína de soja o isoflavonas de soja mejoran la salud ósea tanto en los ratones sanos como en los ratones con huesos osteoporóticos. Este mismo estudio muestra que las isoflavonas de la soja influencian la osteogénesis y la osteoclastogénesis en cultivos celulares, posiblemente a través del mecanismo de unión al receptor de estrógeno (68).
Una revisión publicada por Anderson y Garner en 1998 presentó los resultados de varios laboratorios usando modelos experimentales de ratones hembras OVX, y demostró un efecto beneficioso de los fitoestrógenos sobre la retención de la masa ósea; sin embargo, estos informes apoyan el concepto de que se necesita una dosis umbral de fitoestrógenos por largos periodos de tiempo (meses-años), dependiendo de la especie animal, antes que los resultados sobre la masa y densidad ósea mineral sean observados; adicionalmente, se ha demostrado que dosis extremadamente altas pueden incluso tener efectos adversos sobre el crecimiento óseo (69); razón por la cual las dosis de isoflavonas en cápsulas deben considerarse con cautela debido a la falta de estudios sobre el potencial de estas moléculas .
Zhang y colaboradores publicaron en 2012 su trabajo con ratas prepúberes alimentadas durante 14 días con proteína de soja (PS) y las compararon con otro grupo de ratas prepú-beres tratadas con 17b-estradiol (E2). En este estudio el efecto sobre el hueso fue determinado mediante un análisis de microarreglos de ARN, evaluando la expresión diferencial de genes cuando se suministraba PS, E2 o ambas al mismo tiempo. Los datos de microarreglos indicaron que el efecto de la dieta de soja sobre el hueso tiene algunos puntos en común con E2, sin embargo, el efecto global de los dos tratamientos es diferente, lo cual sugiere nuevas acciones de la dieta de soja sobre el desarrollo óseo. La principal conclusión fue la asociación del efecto osteoblastogenico de las dietas de soja con la activación de la ruta de señalización de proteínas morfogenéticas del hueso (BMP); estas proteínas pertenecen a la familia de los factores de crecimiento transformantes TGF-beta (TGF-p), una superfamilia de proteínas con la capacidad de inducir la formación de hueso nuevo, cartílago y tejido conjuntivo (70).
Estudios in vitro con cultivos de células osteoblásticas. Los osteoblastos son las células directamente involucradas en la formación del hueso y durante su diferenciación ocurren tres procesos determinantes: la proliferación, maduración de la matriz y la mineralización. La actividad osteoblástica puede evidenciarse, entonces, por un incremento en la actividad de ciertos marcadores, como la enzima ALP, la síntesis de colágeno de tipo1 y la presencia de osteopontina.
Choi y colaboradores en 2001 investigaron el efecto de un extracto etanólico de soja sobre la actividad de células MC3T3-E1, una línea celular osteoblástica de la bóveda craneana de ratón, y notaron un aumento en la actividad de la enzima ALP y la síntesis de colágeno en las células que fueron tratadas con extractos de soja en concentración de 0.05 g/L; además, el tratamiento con el fármaco antiestrogénico tamoxifeno eliminó la actividad osteoblástica de las células MC3T3-E1, ocasionada por el extracto de soja, lo cual sugirió que la acción principal del extracto de la soja sobre las células MC3T3-E1 es similar a la del estrógeno, es decir estimula directamente la formación ósea en células osteoblásticas cultivadas in vitro e impidiendo la apoptosis de estas células, efectos que podrían estar asociados con la activación del receptor de estrógeno (71).
Posteriormente Kanno y colaboradores en 2004, publicaron un estudio sobre los efectos de los fitoestrógenos (genisteína, daidzeína y coumestrol) y los estrógenos ambientales [bisfenol A(BPA), p-n-nonilfenol (NP) y bis-(2-etilhexil)ftalato (DEHP)] sobre la diferenciación de osteoblastos, utilizando las células MC3T3-E1 de ratón, y demostraron que el coumestrol en dosis de 10-10 a 10-6 M mejoró ligeramente la proliferación celular; por el contrario, ni los fitoestrógenos (daidzeína, genisteína) ni los estrógenos ambientales aumentaron la proliferación celular; adicio-nalmente, los fitoestrógenos como el cou-mestrol, la genisteína y la daidzeína y el BPA aumentaron los indicadores de la actividad osteoblástica como la ALP y el hueso mejoró la mineralización en las células MC3T3-E1 (72).
Conclusión
El consumo de soja como fuente principal de isoflavonas, a través de la dieta o en forma de suplementos alimenticios, tiene gran importancia por sus efectos beneficiosos para la salud, particularmente, prevención de la osteoporosis, como antioxidante y en la salud cardiovascular.
Aunque se ha demostrado que la potencia transcripcional de las isoflavonas es más débil comparada con la del 17b-estradiol, existe gran aceptación de que el consumo de isoflavonas aumenta la densidad mineral ósea en mujeres postmenopaúsicas y que la disminución en los niveles de estradiol biodisponible en hombres de edad avanzada podría ser la causa principal de la pérdida de masa ósea en esta población.
Sobre el efecto de las isoflavonas en el metabolismo óseo de niños se requieren estudios más profundos que demuestren posibles efectos adversos o beneficiosos.
Investigaciones realizadas en modelos animales y cultivos de líneas celulares osteo-blásticas indican que las isoflavonas de la soja disminuyen la actividad osteoclástica y promueven la diferenciación, proliferación y maduración osteoblástica; actividades que pueden ser mediadas a través de la unión al RE-a y serían dependientes de la concentración de isoflavonas.
Asimismo, estudios recientes realizados en ratones prepúberes presentan fuertes evidencias de la existencia de un mecanismo de acción no estrógenicos involucrado en la activación de la ruta de señalización de proteínas morfogenéticas del hueso (BMP), las cuales tienen la capacidad de inducir la formación de hueso nuevo, cartílago y tejido conjuntivo.
Financiación: Universidad del Atlántico. Barran-quilla (Colombia).
Conflicto de intereses: ninguno.
Referencias
1. Murkies AL, Wilcox G, Davis SR. Clinical review 92: Phytoestrogens. J Clin Endocrinol Metab 1998; 83: 297-303. [ Links ]
2. Ngo Le Van. Coumestrin, a coumestan derivative from soybean roots. Phytochemistry 1984; 23:1204-5. [ Links ]
3. Stechell KD, Cassidy A. Dietary Isoflavones: Biological Effects and Relevance to Human Health. J Nut 1999; 129(3): 758S-67S. [ Links ]
4. Nagata C, Takatsuka N, Shimizu H. Soy product intake and hot flashes in Japanese women: results from a community-based prospective study. Am J Epidemiol 2001;153:790-3. [ Links ]
5. Chi X, Zhang T. The effects of soy isoflavone on bone density in north region of climacteric Chinese women. J Clin Biochem Nutr 2013; 53(2):102-7. [ Links ]
6. Lagari VS, Levis S. Phytoestrogens in the Prevention of Postmenopausal Bone. J Clin Densitom 2013; 16(4):445-9. [ Links ]
7. Tai TY, Tsai KS, Tu ST, Wu JS, Chang CI, Chen CL et al. The effect of soy isoflavone on bone mineral density in postmenopausal Taiwanese women with bone loss: A 2-year randomized double-blind placebo-controlled study. Osteoporos Int 2012; 23(5):1571-80. [ Links ]
8. Alekel DL, Van Loan MD, Koehler KJ, Hanson LN, Stewart JW, Hanson KB et al. The soy isoflavones for reducing bone loss (SIR-BL) study: a 3-y randomized controlled trial in postmenopausal women. Am J Clin Nutr 2010; 91(1): 218-30. [ Links ]
9. Levis S, Strickman-Stein N, Ganjei-Azar P, Xu P, Doerge DR, Krischer J. Soy isoflavones in the prevention of menopausal bone loss and menopausal symptoms: a randomized,Efecto de las isoflavonas de la soja en la salud ósea de adultos y niños double-blind trial. Arch Intern Med 2011;171(15):1363-9. [ Links ]
10. Kang HB, Zhang YE, Yang JD, Lu KL. Study on soy isoflavone consumption and risk of breast cancer and survival. Asian Pac J Cancer Prev 2012; 13(3): 995-8. [ Links ]
11. Mahmoud AM, Yang W, Bosland MC. Soy isoflavones and prostate cancer: a review of molecular mechanisms. J Steroid Biochem Mol Biol 2014; 140:116-32. [ Links ]
12. Zhang Z, Wang CZ, Du GJ, Qi LW, Calway T, He TC et al. Genistein induces G2/M cell cycle arrest and apoptosis via ATM/p53-dependent pathway in human colon cancer cells. Int J Oncol 2013; 43(1):289-96. [ Links ]
13. Hillman GG, Singh-Gupta V. Soy isoflavo-nes sensitize cancer cells to radiotherapy. Free Radic Biol Med 2011; 51(2):289-98. [ Links ]
14. Hallund J, Bügel S, Tholstrup T, Ferrari M, Talbot D, Hall WL et al. Soja isoflavone-en-riched cereal bars affect markers of endote-lial function in postmanopausal women. Br J Nutr 2006; 95(6):1120-6. [ Links ]
15. Hayashi K, Handa K, Koike T, Saito T. The possibility of genistein as a new direct pulpcapping agent. Dent Mater J 2013; 32(6):976-85. [ Links ]
16. Bhathena SJ, Velasquez MT. Beneficial role of dietary phytoestrogens in obesity and diabetes. Am J clin Nutr 2001; 76(6):1191-201. [ Links ]
17. Anderson JW. Beneficial effects of soy protein consumption of renal function. Asian Pac J Clin utr 2008;17 (Supl1): 324-8. [ Links ]
18. Henry-Vitrac C, Berbille H, Mérillon JM, Vi-trac X. Soy isoflavones as potential inhibitors of Alzheimer β-amyloid fibril aggregation in vitro. Food Research International 2010;43 (8): 2176-8. [ Links ]
19. Luo S1, Lan T, Liao W, Zhao M, Yang H.Genistein inhibits Aß25-35 -induced neurotoxicity in PC12 cells via PKC Signaling Pathway. Neurochem Res 2012; 37(12):2787-94. [ Links ]
20. Winkel-Shirley B. Flavonoid biosynthesis. A colorful model for genetics, biochemistry, cell biology, and biotechnology. Plant Phy-siol 2001; 126: 485-93. [ Links ]
21. Setchell KDR. Phytoestrogens: the biochemistry, and implications for human health of soy isoflavones. Am J Clin Nutr 1998; 68:1333S-46S. [ Links ]
22. Mitchell JH, Gardner PT, McPhail DB, Morrice PC, Collins AR, Duthie GG. Antioxi-dant efficacy of phytoestrogens in chemical and biological model systems. Arch Biochem Biophys 1998; 360(1): 142-8. [ Links ]
23. Pailo A, Mota-Filipe H. Effects of some natural 5-hydroxy-isoflavones on cultured human endothelial cells in presence and absence of hydrogen peroxide. J Pharm Pharmacol 2006; 58(1):101-5). [ Links ]
24. Mazur W, Duke J, Wahala K, Rasku S, Adlercreutz H. Isoflavonoids and lignans in legumes: nutritional and health aspects in humans. J Nutr Biochem 1998; 9: 193-200. [ Links ]
25. Manzur W, Adlercreutz H. Natural and anthropogenic environmental oestrogens: the scientific basis for risk assessment. Pure Appl Chem 1998; 70: 1759-76. [ Links ]
26. Ogbuewu IP, Uchegbu MC, Emenalom OO, Okoli IC, Iloeje MU. Overview of the chemistry of soy isoflavones, potential threats and potential therapeutic benefits. EJEAFChe 2010; 9(4): 682-95. [ Links ]
27. Wang HJ, Murphy PA. Isoflavone content in commercial soybean foods. J Agric Food Chem 1994; 42: 1666-73. [ Links ]
28. Murphy PA, Song T, Buseman G, Barua K, Beecher GR, Trainer D, et al. Isoflavones in retail and institutional soy foods. J Agric Food Chem 1999; 47:2697-2704. [ Links ]
29. Barnes S. The biochemistry, chemistry and physiology of the isoflavones in soybeans and their food products. Lymphat Res Biol 2010 Mar;8(1):89-98. [ Links ]
30. Day AJ, Canada FJ, Diaz JC, Kroon PA, Mclauchlan R, Faulds CB, et al. Dietary fla-vonoid and isoflavone glycosides are hy-drolysed by the lactase site of lactase phlorizin hydrolase. FEBS Lett 2000; 468:166-70. [ Links ]
31. Muthyala RS, Ju YH, Sheng S, Williams LD, Doerge DR, Katzenellenbogen BS, et al. Equol, a natural estrogenic metabolite from soy isoflavones: convenient preparation and resolution of R- and S-equols and their differing binding and biological activity through estrogen receptors alpha and beta. Bioorg Med Chem 2004; 12:1559-67. [ Links ]
32. Setchell KD, Clerici C, Lephart ED, Cole SJ, Heenan C, Castellani D et al. S-equol, a potent ligand for estrogen receptor beta, is the exclusive enantiomeric form of the soy isoflavone metabolite produced by human intestinal bacterial flora. Am J Clin Nutr 2005;81:1072-9. [ Links ]
33. Tousen Y, Ezaki J, Fujii Y, Ueno T, Nishimuta M, Ishimi Y. Natural S-equol decreases bone resorption in postmenopausal, non-equol-producing Japanese women: A pilot randomized, placebo-controlled trial. Menopause 2011; 18(5):563-74. [ Links ]
34. Ronis MJ, Little JM, Barone GW, Chen G, Radominska-Pandya A, Badger TM. Sulfation of the isoflavones genisteina and daid-zein in human and rat liver and gastrointestinal tract. J Med Food 2006;9:348-55. [ Links ]
35. Setchell KD, Brown NM, Desai P, Zimmer-Nechemias L, Wolfe BE, Brashear WT et al. Bioavailability of pure isoflavones in healthy humans and analysis of commercial soy isoflavone supplements. J Nutr 2001;131:1362S-75S. [ Links ]
36. Rozman K, Bhatia Calafat AM, Chambers C, Culty M, Etzel RA, Flaws JA et al. NTP-CERHR Expert Panel Report on the Reproductive and Developmental Toxicity of Soy Formula. Birth Defects Res B Dev Reprod Toxicol 2006; 77(4): 280-397. [ Links ]
37. Barnes S, Prasain J, D'Alessandro T, Arabsh-ahi A, Botting N, Lila MA et al. The metabolism and analysis of isoflavones and other dietary polyphenols in foods and biological systems. Food Funct 2011; 2(5):235-44. [ Links ]
38. Sadowska-Bartosz I, Adamczyk R, Bartosz G. Protection against peroxynitrite reactions by flavonoids. Food Chem 2014; 164:228-33. [ Links ]
39. Boersma BJ, D'Alessandro T, Benton MR,Kirk M, Wilson LS, Bottinh NP, et al. Neutrophil myeloperoixidase chlorinates and nitrates soy isoflavones and enhances their antioxidant properties. Free Radic Biol Med 2003; 35 (11): 1417-30. [ Links ]
40. Setchell KD. Soy isoflavones-benefits and risks from nature's selective estrogen receptor modulators (MSREs). J Am Coll Nutr 2001; 20(5 Suppl):354S-362S. [ Links ]
41. Parfitt AM. Bone remodeling and bone loss: understanding the pathophysiology of osteoporosis. Clin Obstet Gynecol 1987; 30(4):789-811. [ Links ]
42. Rodan GA. Introduction to bone biology. Bone 1992; 13(Supl 1):S3-6. [ Links ]
43. Korach KS, Migliaccio S, Davis VL. Estrogens. In: Munson PL (ed). Principles of Pharmacology-Basic Concepts and Clinical Applications. New York: Chapman and Hall; 1994. p. 89-25. [ Links ]
44. Jensen EV. Steroid hormones, receptors and antagonists. Ann NY Acad Sci 1995; 761:1176. [ Links ]
45. Beato M, Herrlich P, Schutz G. Steroid hormone receptors: many actorsin search of a plot 1995; 83:851-7. [ Links ]
46. Tsai M-J, O'Malley BW. Molecular mechanisms of action of steroid/ thyroid receptor superfamily members. Annu Rev Biochem 1995; 63;451-86. [ Links ]
47. Nakamura T, Imai Y, Matsumoto T, Sato S, Takeuchi K, Igarashi K et al. Estrogen Prevents Bone Loss via Estrogen Receptor a and Induction of Fas Ligand in Osteoclasts.Cell 2007; 130: 811-23. [ Links ]
48. García AJ, Tom C, Guemes M, Polanco G, Mayorga ME, Wend K et al. ERa signaling regulates MMP3 expression to induce FasL cleavage and osteoclast apoptosis. J Bone Miner Res 2013; 28(2):283-90. [ Links ]
49. Okazaki R, Inoue D, Shibata M, Saika M, Kido S, Ooka H et al. Estrogen promotes early osteoblast differentiation and inhibits adipocyte differentiation in mouse bone marrow stromal cell lines that express estrogen receptor (ER) alpha or beta. Endocrinology 2002; 143:2349-56. [ Links ]
50. Cao L, Bu R, Oakley JI, Kalla SE, Blair HC.Estrogen receptor-beta modulates synthesis of bone matrix proteins in human osteo-blast-like MG63 cells. J Cell Biochem 2003; 89(1):152-64. [ Links ]
51. Almeida M, Iyer S, Martin-Millan M, Bar-tell SM, Han L, Ambrogini E et al. Estrogen receptor-a signaling in osteoblast progenitors stimulates cortical bone accrual. J Clin Invest 2013; 123(1):394-404. [ Links ]
52. Cutler GB Jr. The role of estrogen in bone growth and maturation during childhood and adolescence. J Steroid Biochem Mol Biol 1997; 61(3-6):141-4. [ Links ]
53. Bilezikian JP, Morishima A, Bell J, Grumbach MM. Increased bone mass as a result of estrogen therapy in a man with aromata-se deficiency. N Engl J Med 1998; 339(9):599-603. [ Links ]
54. Yilmaz D, Ersoy B, Bilgin E, Gümüser G, Onur E, Pinar ED. Bone mineral density in girls and boys at different pubertal stages: relation with gonadal steroids, bone formation markers, and growth parameters. J Bone Miner Metab 2005; 23:476-82. [ Links ]
55. Doneray H, Orbak Z. Association between anthropometric hormonal measurements and bone mineral density in puberty and constitutional delay of growth and puberty. West Indian Med J 2010; 59(2):125-30. [ Links ]
56. Khosla S, Melton LJ, Atkinson EJ, O'Fallon WM. Relationship of serum sex steroid levels to longitudinal changes in bone density in young versus elderly men. J Clin Endocrinol Metab 2001; 86(8):3555-61. [ Links ]
57. Mac Gregor JI, Jordan VC. Basic guide to the mechanisms of antiestrogen action. Pharmacol Rev 1998; 50: 151-96. [ Links ]
58. Pons DG, Nadal-Serrano M, Del Mar Blan-quer-Rossello M, Sastre-Serra J, Oliver J, Roca P. Genistein Modulates Proliferation and Mitochondrial Functionality in Breast Cancer Cells Depending on ERalpha/ERbeta Ratio. J Cell Biochem 2014; 115(5):949-58. [ Links ]
59. Jefferson WN, Newbold RR. Potential endocrine-modulating effects of various phytoestrogens in the diet. Nutrition 2000; 16:658-62. [ Links ]
60. Kostelac D, Rechkemmer G, Briviba K. Phytoestrogens modulate binding response of estrogen receptors alpha and beta to the estrogen response element. J Agric Food Chem 2003;51(26):7632-5. [ Links ]
61. Setchell KD, Zimmer-Nechemias L, Cai J, Heubi JE. Exposure of infants to phyto-oestrogens from soy-based infant formula. Lancet 1997; 350(9070):23-7. [ Links ]
62. Giampietro PG, Bruno G, Furcolo G, Casati A, Brunetti E, Spadoni GL et al. Soy protein formulas in children: no hormonal effects in long-term feeding. J Pediatr Endocrinol Metab 2004; 17(2):191-6. [ Links ]
63. Cao Y, Calafat AM, Doerge DR, Umbach DM, Bernbaum JC, Twaddle NC et al. Isofla-vones in urine, saliva, and blood of infants: data from a pilot study on the estrogenic activity of soy formula. J Expo Sci Environ Epidemiol 2009; 19(2):223-34. [ Links ]
64. Khalil D, Lucas E, Juma S, Smith B, Payton M, Arjmandi B. Soy Protein Supplementation Increases Serum Insulin-Like Growth Factor-I in Young and Old Men but Does Not Affect Markers of Bone Metabolism. J Nutr 2002; 132(9):2605-8. [ Links ]
65. Ishimi Y, Miyaura C, Ohmura M, Onoe Y, Sato T, Uchiyama Y et al. Selective effects of genistein, a soybean isoflavone, on B-lymphopoiesis and bone loss caused by estrogen deficiency. Endocrinology 1999;140(4):1893-900. [ Links ]
66. Ishimi Y, Arai N, Wang X, Wu J, Umegaki K, Miyaura C et al. Difference in effective dosage of genistein on bone and uterus in ovariectomized mice. Biochem Biophys Res Commun 2000; 274(3):697-701. [ Links ]
67. Fujioka M, Sudo Y, Okumura M, Wu J, Ue-hara M, Takeda K et al. Differential effects of isoflavones on bone formation in growing male and female mice. Metabolism 2007;56(8):1142-8. [ Links ]
68. Chin KY, Ima-Nirwana S. Can soy prevent male osteoporosis? a review of the current evidence. Curr Drug Targets 2013;14(14):1632-41. [ Links ]
69. Anderson JB, Garner SC. Phytoestrogens and bone. Baillikre's Clinical Endocrinologyand Metabolism 1998; 12(4): 543-57. [ Links ]
70. Zhang J, Lazarenko OP, Wu X, Tong Y, Blackburn ML, Gomez-Acevedo H et al. Differential effects of short term feeding of a soy protein isolate diet and estrogen treatment on bone in the pre-pubertal rat. PLoS One 2012; 7(4):e35736. [ Links ]
71. Choi EM, Suh KS, Kim YS, Choue RW, Koo SJ. Soybean ethanol extract increases the function of osteoblastic MC3T3-E1 cells. Phytochemistry 2001; 56(7):733-9. [ Links ]
72. Kanno S, Hirano S, Kayamab F. Effects of phytoestrogens and environmental estrogens on osteoblastic differentiation in MC3T3-E1 cells. Toxicology 2004; 196: 137-45. [ Links ]
