










Services on Demand
Journal
Article
 Spanish (pdf)
Spanish (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mailIndicators
-
 Cited by SciELO
Cited by SciELO -
 Access statistics
Access statistics
Related links
-
 Cited by Google
Cited by Google -
 Similars in
SciELO
Similars in
SciELO -
 Similars in Google
Similars in Google
Share

 Permalink
PermalinkRevista Salud Uninorte
Print version ISSN 0120-5552On-line version ISSN 2011-7531
Salud, Barranquilla vol.31 no.3 Barranquilla Sep./Dec. 2015
https://doi.org/10.14482/sun.31.3.567
http://dx.doi.org/10.14482/sun.31.3.567
Diversidad de roedores, hantavirus y su relación con la salud pública
Diversity of rodents, hantavirus and its relationship to public health
Camilo Guzmán1, Salim Mattar2, Alfonso Calderón3
1 QF. M. Sc. Instituto de Investigaciones Biológicas del Trópico (IIBT). Profesor asociado, Programa Regencia en Farmacia, Facultad Ciencias de la Salud, Universidad de Córdoba, Córdoba. Doctorante en Medicina Tropical SUE-Caribe. cguzman40@hotmail.com
2 Ph. D. Instituto de Investigaciones Biológicas del Trópico (IIBT). Profesor titular, Facultad Medicina Veterinaria y Zootecnia, Universidad de Córdoba, Córdoba. mattarsalim@hotmail.com
3 MVZ, M. Sc Instituto de Investigaciones Biológicas del Trópico (IIBT). Profesor titular, Facultad Medicina Veterinaria y Zootecnia, Universidad de Córdoba, Córdoba. Doctorante en Medicina Tropical SUE-Caribe. acalderonr@correo.unicordoba.edu.co
Correspondencia: Salim Mattar, Instituto de Investigaciones Biológicas del Trópico (IIBT). mattarsalim@hotmail.com, smattar@correo.unicordoba.edu.co
Fecha de recepción: 10 de junio de 2015
Fecha de aceptación: 17 de julio de 2015
Resumen
El orden Rodentia comprende el grupo de mamíferos con la mayor diversidad de especies vivientes que son hospederos y reservorios naturales de hantavirus. Los hantavirus que causan síndrome cardiopulmonar en América utilizan como reservorio a la subfamilia Sigmodontinae. Cada hantavirus posee afinidad con un roedor hospedero específico. En 1976 en Corea se cultivó del pulmón del ratón Apodemus agrarius, el primer virus Hantaan. En 1978 se identificó el virus Seoul de Rattus norvegicus. En 1997 en Finlandia se aisló el hantavirus Puumala del pulmón de Myodes glareolus. En 1993 se aisló hantavirus Dobrava (Eslovenia) de Apodemus flavicolis y en 1982 fue aislado de Microtus pennsylvanicus el primer hantavirus no patógeno en las Américas: Prospect Hill. No obstante, en 1993 una misteriosa enfermedad pulmonar se expandió en Four Corners (USA); los pacientes repentinamente enfermaron y muchos murieron debido a shock y/o edema pulmonar; los síntomas no fueron asociados con hantavirus. Más tarde se aisló un nuevo hantavirus del roedor Peromyscus maniculatis, el cual era desconocido, y se lo denominó como Virus Sin Nombre y se asoció como la causa de este brote epidémico. En Suramérica se han reportado hantavirus en distintos hospederos naturales. En Colombia, entre 2004 y 2015 se han publicado diversos estudios realizados en el Caribe colombiano y en Urabá en los que se evidenció la presencia de hantavirus en humanos y en roedores. Recientemente se tuvo el primer reporte serológico de infección por hantavirus en humanos en la región de la Orinoquia colombiana. Sin embargo, la hantavirosis no es considerada una enfermedad de notificación obligatoria, y es probable que actualmente esté en silencio epidemiológico.
Palabras clave: Bunyaviridae, epidemias, noxas, reservorios, roedores, virus (MeSH).
Abstract
The order Rodentia comprises the group of mammals with the greatest diversity of living species that are natural hosts and reservoirs of hantavirus. Guests are natural reservoirs and hantavirus. All hantavirus that cause cardiopulmonary syndrome in the Americas are linked to the subfamily Sigmodontinae. Each hantavirus has affinity for a specific rodent host. In Korea the first Hantaan virus was isolated in 1976 from a lung culture of field mouse Apodemus agrarius. In 1978, the Seoul virus was identified from Rattus norvegicus. In Finland, the Puumala hantavirus from Myodes glareolus lung. In 1993, hantavirus Dobrava from Apodemus flavicolis was isolated in Slovenia and in 1982 the first hantavirus in the Americas called Prospect Hill isolated form Microtus pennsylvanicus, which resulted nonpathogenic and for this reason it was believed that the American hantavirus were nonpathogenic. However, in 1993 a mysterious lung disease began spreading in Four Corners (USA), the patients suddenly became ill and many died due to shock and/or lung edema; these early symptoms were not associated with hantavirus but a virus subsequently isolated from Peromyscus maniculatis was a new virus now known as hantavirus without name was cause of this outbreak. In South America hantavirus have been reported in different natural hosts. In Colombia between 2004 and 2006 the first serological studies in the Colombian Caribbean, where they reported the circulation of hantavirus in humans and rodents, were published. However, hantavirus is not considered a reportable disease and probably currently it is in epidemiological silence.
Keywords: Bunyaviridae, epidemics, host, noxae, rodentia, viruses. (MeSH).
DIVERSIDAD DE ROEDORES, HANTAVIRUS Y SU RELACIÓN CON LA SALUD PÚBLICA
El orden Rodentia comprende un grupo de mamíferos con la mayor diversidad de especies vivientes en el mundo, la más amplia gama de formas adaptativas y el mayor número de hábitats conocidos. Poseen altas tasas de evolución, periodos cortos de vida, múltiples camadas y crías numerosas (1). Son aproximadamente 2300 especies silvestres; más del 40 % de estos mamíferos pertenecen a este orden; los roedores son encontrados en todos los suelos y ocupan casi todos los tipos de hábitat disponibles, excepto la región polar (1). Los roedores múridos (orden Rodentia, familia Muridae) son los hospederos y reservorios naturales de los hantavirus. Los hantavirus son virus emergentes que causan fiebres hemorrágicas con síndrome renal (FHSR) en Asia y Europa. La infección por hantavirus en las Américas causa síndrome cardiopulmonar por hantavirus (SCPH). Los estudios de fósiles aportan pruebas de la presencia de roedores múridos en los últimos 20 millones de años en América del Norte y en los últimos 3,5 millones de años en América del Sur (2,3). Sus hábitos son frecuentemente nocturnos. En regiones cálidas como el Caribe, la procreación puede producirse en forma ininterrumpida durante todo el año. Es probable que la mayoría de los individuos vivan menos de dos años; sin embargo, el enorme potencial reproductivo de algunas especies hace que la población aumente en forma exponencial; después de ello sigue una disminución repentina del número de animales cuando se agota el alimento en una zona particular. Estas fluctuaciones pueden mostrar una periodicidad de tres a cuatro años en algunas especies y hábitats (4).
Todos los hantavirus que causan SCPH en el continente americano están ligados a la subfamilia Sigmodontinae, que agrupa a más de 420 especies vivientes (5,6), habitan en entornos rurales, aunque algunos no tienen predilección por un hábitat particular (3).
Cada hantavirus posee afinidad por un roedor hospedero específico (7,8); este hecho de estar dentro de una especie predominante de animales restringe la distribución de otros virus. La distribución del virus puede observarse en hospederos diversos, o bien limitarse a miembros muy selectos de la especie de roedores. La existencia de niveles altos de concordancia entre el hospedero y la filogenia de hantavirus refuerza la relación muy duradera y quizá de evolución compartida (coevolución) entre el virus y su hospedero.
Familia Muridae. Son llamados comúnmente ratas y ratones, silvestres y domésticos; se caracterizan en general por su tamaño pequeño a mediano (<300g), tienen el pelaje suave o ligeramente erizado (un solo un género es semiespinoso), los ojos pequeños o medianos y las orejas medianas y generalmente sobresalientes. Las extremidades son siempre cortas y delicadas, y los dedos poseen garras curvas, el tamaño de la cola es muy variables, desde cortas a largas. Entre las especies terrestres domina la cola corta y las patas largas y estrechas, mientras que en las arbóreas la cola es larga y las patas cortas y anchas. Un reducido número de especies presenta adaptaciones semiacuáticas y semicavadoras, con desarrollo de membranas interdigitales o peines laterales, y rostros y garras alargadas, respectivamente (4, 6).
Se conoce que la familia Muridae desde el Eoceno habitaba en América del Norte y Eurasia, se diversificó notoriamente durante el Mioceno, hace unos 20 millones de años (Ma), y en la actualidad ha alcanzado la mayor diversidad de los mamíferos vivientes en todo el mundo. De esta familia se conoce solamente que los representantes de la subfamilia Sigmodontinae ingresaron a Sudamérica durante el Montehermosense (hace unos 3.5 Ma), diversificándose el grupo previamente durante el Mioceno tardío y el Plioceno de América del Norte (mayormente durante el henfiliense y el blanquenc, hace unos 4 Ma). A partir del gran intercambio de fauna americano con el istmo de Panamá los sigmodontinos que ingresaron a Suramérica se dispersaron y se adaptaron durante los últimos 3 Ma (2,3).
Actualmente los sigmodontinos agrupan a más de 420 especies vivientes, lo cual demuestra su alta capacidad adaptativa. Esta rápida colonización en América ha sido posible gracias a sus atributos biológicos: un periodo corto de vida, gran variedad de tipos y estrategias alimentarias y de locomoción, y una gran capacidad de dispersión. Estos atributos biológicos han sido también favorecidos por otros dos factores históricos: la aparente ausencia de roedores nativos de tamaño pequeño y la gran heterogeneidad y dinámica de los ambientes naturales durante el Neógeno y el Cuaternario sudamericano (2, 3).
Por otro lado, es interesante señalar otra inmigración más reciente de roedores múridos a Sudamérica, ocurrida con la llegada de los europeos a partir de 1492, con la introducción de ratas y ratones domésticos (Rattus y Mus). La agresividad y rápida expansión de estos roedores ha causado cambios notables en la composición y estructura de muchas faunas locales de micromamíferos residentes (1, 4, 6).
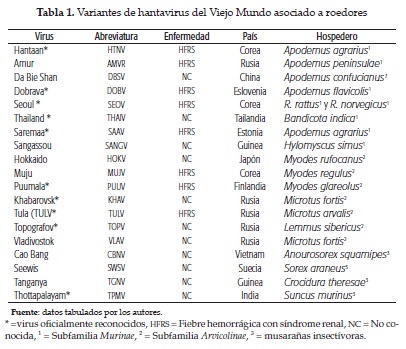
Asociación de los sigmondotinos y hantavirus. Los roedores de la subfamilia Sigmodontinae de múridos que han sido considerados hospederos de los virus que causan síndrome cardiopulmonar por hantavirus (SCPH) viven bien en entornos rurales, aunque algunos no tienen predilección por un hábitat particular. La preferencia de los roedores a entrar en las viviendas y edificios vecinos es importante en salud pública (9,10,11, 12). Esta característica del ratón de campo fue un factor básico en la epidemia de hantavirus en 1993 en el suroeste de los Estados Unidos. Otros virus detectados a menudo en América del Norte se hospedan en miembros de la subfamilia Arvicolinae, pero al parecer no causan enfermedad en los humanos. Los hantavirus del Viejo Mundo que causan fiebre hemorrágica con síndrome renal (FHSR) utilizan como reservorios algunos miembros de la subfamilia Arvicolinae (10). Algunos roedores comunes que viven en asociación con las personas, como el ratón casero M. Musculus, pertenecen a otras subfamilias y al parecer no son reservorios importantes de hantavirus, pero sí para otros microorganismos, como los arenavirus (13,14,15).
El descubrimiento de un nuevo agente zoonótico. Las enfermedades infecciosas son a menudo conocidas desde mucho antes del descubrimiento de los agentes microbiológicos que las causan; este es el caso de los hantavirus, cuyos síntomas, similares a los que se conocen hoy día como fiebre hemorrágica con síndrome renal por hantavirus (FHSR), fueron descritos en China hace más de 1000 años (16). Enfermedades como la fiebre epidémica hemorrágica y fiebre Songo captaron la atención durante el siglo XX, asociadas a conflictos humanos; ejemplo, la nefritis de las trincheras, que afectó a soldados durante la Primera Guerra Mundial (17). Durante la guerra de Corea, entre 1950-1953, más de 3000 soldados americanos padecieron enfermedades y muchos murieron de una enfermedad asociada con falla renal y algunas veces con manifestaciones hemorrágicas severas (18). Esta enfermedad fue llamada 'fiebre hemorrágica coreana' y se sugirió de forma equivocada que era producida por un vector artrópodo portador del agente etiológico y no se percibió a los ratones como el verdadero hospedero del virus hanta (19).
El descubrimiento se produjo solo en 1976 cuando Ho Wang Lee et al. (20) aislaron el virus Hantaan del Apodemus agrarius, y fue llamado así por su cercanía al río de su nombre (6). Pocos años después del descubrimiento del virus fue propagado en células carcinogénicas humanas (A-549), y la observación al microscopio electrónico demostró un nuevo género de la familia Bunyaviridae (21).
Cronología de la infección por hantavirus en el Viejo Mundo. Posterior al descubrimiento del virus Hantaan, otro hantavirus patogénico coreano fue identificado: el virus Seoul del Rattus norvegicus (22). En el sureste de Finlandia, del pulmón de Myodes glareolus se aisló el hantavirus Puumala, llamado así por el área donde fue capturado el roedor, y parece ser el agente causal de la nefropatía epidémica (NE), una variante menos agresiva de FHSR (23,24), la cual fue descrita en el país vecino de Suecia en 1934 (25). En 1993 se aisló un hantavirus de Apodemus flavicolis en Eslovenia y se le llamó virus Dobrava (26).
Cronología de la infección por hantavirus en el Nuevo Mundo. En 1982 el primer hantavirus reportado en las Américas fue el Prospect Hill (27,28), aislado de Microtus pennsylvanicus; dado que este virus es apatogéno, se creyó que los hantavirus americanos no eran patógenos. En 1993 una misteriosa enfermedad pulmonar comenzó a expandirse en Four Corners (USA), donde confluyen los estados de Nuevo México, Colorado, Utah y Arizona. Los pacientes repentinamente enfermaron y muchos murieron debido a shock y/o edema pulmonar; estos síntomas no fueron asociados en ese momento con hantavirus; posteriormente se aisló del Peromyscus maniculatis, un nuevo hantavirus conocido actualmente como 'virus sin nombre', causante de este brote epidémico (29).
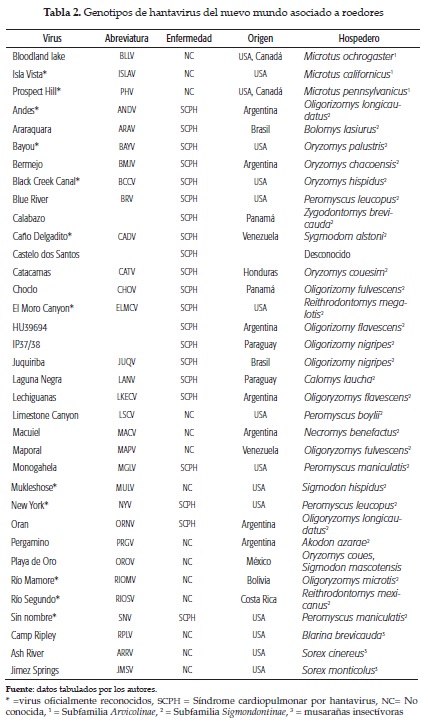
Los hantavirus en Latinoamérica. En Argentina se describió la fiebre hemorrágica argentina en 1953 como una nueva enfermedad, y en 1958 se logró aislar el virus Junín (arenavirus) como agente etiológico de FHA, siendo este el primer país latinoamericano donde más se profundizó en el conocimiento de las fiebres hemorrágicas. En 1996, después de un ataque severo de síndrome cardiopulmonar, se aisló un hantavirus del roedor Oligoryzomys longicaudatus y se le llamó virus Andes. Desde entonces ha habido una explosión de hantavirus en Suramérica, con más de 15 nuevas variantes reportadas hasta ahora (figura 1). Cerca de 54 diferentes variantes de hantavirus han sido identificadas alrededor del mundo (tablas 1 y 2); algunos representan oficialmente nuevos hantavirus reconocidos, mientras que otros son nuevas cepas de virus ya conocidos. La notable similitud entre la evolución genealógica de los hantavirus y de sus roedores hospederos demuestra millones de años de coevolución (30), desde un ancestro común de roedores (Muridae) subfamilias , Murinae, Sigmodontinae y Arvicolinae, probablemente desde hace 50 millones de años (31).
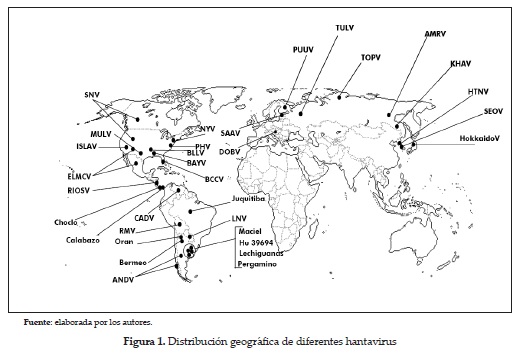
SAAV, DOBV, PUUV, TULV, TOPV, AMRV, KHAV, HTNV, SEOV, Hokkaido V, SNV, MULV, ISLAV, NYV, PHV, BLLV, BAYV, BCCV, ELMVC, RIOSV, Choclo, Calabazo, CADV, RMV, Oran, Bermeo, ANDV, Juanquitiba, LNV, Maciel (Hu 39694, Lechiguanas, pergamino) (tabla 1) .
Reservorios y relaciones genéticas entre el virus y su hospedero. La serología y las relaciones genéticas entre los varios tipos de hantavirus son paralelas a las relaciones genéticas entre los hospederos roedores predominantemente para cada virus. Este paralelismo entre las relaciones genéticas de los hantavirus y sus hospederos roedores fortalece la hipótesis de que los hantavirus no emergieron debido a mutaciones recientes, sino que aparecieron debido a disturbios ecológicos que condujeron a roedores infectados con hantavirus a estar en contacto más estrecho con el hombre. Los datos filogenéticos, además de la correlación geográfica y clínica, muestran que los hantavirus tienen una asociación con sus hospederos, quizás debido a una evolución con sus hospederos roedores específicos (30, 31).
Características biológicas de los reservorios Roedentia. El comportamiento del hospedero puede tener una importante influencia en la transmisión promedio de un patógeno (32). Experimentos de laboratorio con el virus Seoul (SEOV) demostraron que los hantavirus son más efectivamente transmitidos por inyección vía intramuscular que por inhalación de aerosoles (33); por otro lado, una correlación entre la apariencia de las heridas y la infección de hantavirus en roedores silvestres ha sido observada para SEOV y SNV (34, 35). Estos hallazgos sugieren que un mayor comportamiento agresivo del huésped es la principal ruta de transmisión de estos virus entre los reservorios.
Sexo. Los machos son más susceptibles a infecciones por hantavirus que las hembras, debido a las heridas que sufren en peleas con otros machos (36), y se convierten en reservorios de hantavirus (37); esto ocurre después de la maduración sexual, y la actividad hormonal en los machos ha sido la hipótesis de la diferencia entre los sexos (38). Algunas hormonas podrían generar cambios en el comportamiento del macho, y además incrementar su exposición para los virus o, alternativamente, regular las hormonas de la respuesta inmune, en relación con los machos más susceptibles para la infección (39) o una combinación de ambos. Cuando las ratas adultas fueron inoculadas experimentalmente con virus SEOV, ambos sexos fueron susceptibles a la infección, sin embargo, los machos esparcieron el virus más tiempo y por muchas rutas, y se presentaron altos niveles de ARN viral en sangre (40, 41).
Edad. En general, la infección por hantavirus no es influenciada por la edad y la infección de animales es crónica y no patogénica (42). Experimentalmente se ha corroborado la transmisión horizontal entre los roedores por medio de aerosoles infecciosos (43, 44, 45). En el campo, la seroprevalencia entre los roedores suele aumentar con el peso corporal y, por consiguiente, con la edad, lo cual destaca la importancia predominante de la transmisión horizontal en la conservación del virus dentro de poblaciones del reservorio (46). Las crías de hembras infectadas muestran anticuerpos maternos circulantes, pero no existen pruebas definitivas de que se produzca una transmisión viral de tipo vertical. De este modo, la persistencia de los hantavirus en los roedores que actúan como reservorios ocurre principalmente durante encuentros intraespecíficos después del destete (47).
Persistencia viral y respuesta inmune en roedores. La infección de hantavirus en humanos es transitoria y reconocida por la respuesta inmune, mientras que la infección en los hospederos naturales se cree que es persistente. La razón detrás de la persistencia del virus puede ser debido a la habilidad de algunos hantavirus después de la infección para bajar la regulación de la replicación o porque induzcan variación antigénica (48). La regulación de las células T ha sido sugerida para mediar la persistencia de SEOV en ratas noruegas (49) y la persistencia de SNV en el ratón ciervo (50). La asociación de hantavirus a tejidos grasosos y oscuros de los hospederos naturales ha sido sugerida en la importancia del mantenimiento (51). Sin embargo, esto ha sido debatido debido a que en el estado natural la persistencia viral por pocos meses es probablemente suficiente para abarcar toda la vida del hospedero (48).
Densidad de la población reservorio e infección en humanos. Muchas poblaciones de roedores muestran variaciones cíclicas en la densidad con periodos de recurrencia de pocos animales; la densidad de la población huésped de roedores está conectada al número de infecciones de hantavirus entre humanos (34, 37). Un alto número de roedores resulta en un alto número de roedores infectados. Un ejemplo es el incremento en la población de roedores que generó el brote de SCPH en Estados Unidos en 1993, el cual se relacionó con un incremento en la precipitación, asociado con el Fenómeno de El Niño, que produjo un incremento de las fuentes de alimento (52). Sin embargo, la correlación de la seroprevalencia con la alta densidad de hospederos no está completamente clara, y esta aparente relación podría estar afectada de la frecuencia de muestreo en los estudios de campo. La densidad de la población hospedera puede ser inversamente proporcional a la seroprevalencia de hantavirus, como lo demostraron varios estudios epidemiológicos en las Américas (50,53). Este fenómeno podría ser explicado porque en realidad la alta densidad de roedores con altos sucesos reproductivos de la población adulta genera una población con alta proporción de animales jóvenes seronegativos (46). Cuando esta población decrece se incrementa la seroprevalencia, debido a que gradualmente aumenta la proporción de animales viejos infectados (54). Sin embargo, aunque la seroprevalencia es baja durante periodos de alta densidad de roedores, puede haber un riesgo silencioso para infección de humanos (46).
Impacto ambiental e infección por hantavirus. La densidad de la población de roedores está muy influenciada por las variaciones de muchos factores ambientales, tales como tipo de vegetación, provisión de alimentos, abundancia de predadores, de nieve y altas temperatura. Fotografías satelitales junto con sistemas de información geográfica considerando el tipo de vegetación de áreas, elevación, declives y características hidrológicas han sido usados para estimar el estatus de infección de ratón ciervo, con el 80 % de precisión y, subsecuentemente, también el riesgo de infección humana (55). Las fuertes lluvias posteriores al Fenómeno de El Niño en 1993 causaron un aumento de la oferta de alimentos de origen vegetal, lo cual produjo un incremento de las poblaciones de roedores y la subsiguiente transmisión de la infección de hantavirus a humanos en los Estados Unidos.
Transmisión. Todos los géneros de Buyanviridae, incluido hantavirus, por lo general se transmiten por contacto con excretas de roedores. El alto número de roedores infectados origina alto riesgo de transmisión a humanos, lo cual significa que muchos factores que influencian la transmisión de hantavirus entre roedores indirectamente también influencian el riesgo de que los humanos adquieran la infección (34).
Transmisión humano a humano. El virus Andes es el único hantavirus conocido y documentado que puede trasmitirse de persona a persona. Los primeros reportes de transmisión interpersonal sospechada se hicieron en Argentina y Chile entre 1996-1998, y estuvieron basados en datos epidemiológicos; en dichos países fueron estudiados grupos de casos para encontrar la probable fuente de la infección (56). Padula et al. (57) encontraron la primera evidencia molecular de transmisión persona a persona. En ese estudio, una serie de 16 pacientes aparentemente relacionados epidemiológicamente estaban infectados con ANDV con una secuencia nucleotídica idéntica; una explicación hipotética pudo ser que los pacientes fueron infectados con una cepa genéticamente idéntica de hantavirus, algunos de idénticos roedores o procedentes de roedores de la misma población local.
Sin embargo, dada la larga distancia entre la residencia de los pacientes, algunas veces superior a 1400 km, la explicación más razonable es que ellos fueron infectados a través de la transmisión persona a persona mientras estaban hospitalizados (58). El hallazgo de la transmisión interpersonal de ANDV puede estar limitado a un corto periodo en la última o en la fase temprana cardiopulmonar, y probablemente demanda el cierre del contacto con una persona infectada (59). Aparte del brote de 1996 en Argentina, la infección de ANDV en trabajadores hospitalarios, la seroprevalencia entre los trabajadores sanitarios no es diferente de la observada en la población general (58).
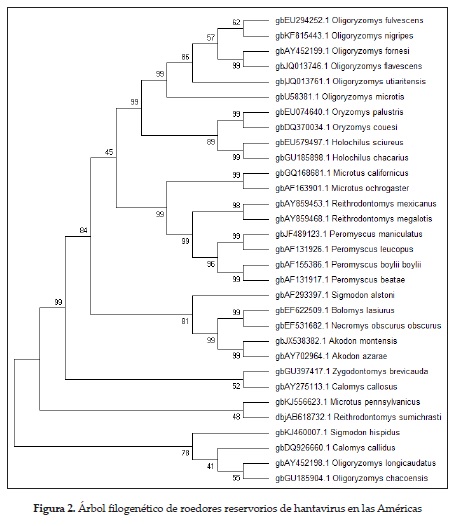
En Suramérica se ha descrito la presencia de hantavirus en distintos hospederos naturales (figura 2); los países limítrofes con Colombia tienen plenamente identificados los reservorios y caracterizadas genotípicamente las cepas patógenas y no patógenas de hantavirus; en Panamá, Oligoryzomyz fulvescens es el reservorio natural del hantavirus choclo, el cual ha sido asociado con casos de SCPH; en Venezuela, Sigmodon alstoni es reservorio del hantavirus caño delgadito, el cual no ha sido asociado a infección en humanos; los estudios en roedores de Bolivia identificaron por primera vez un hantavirus propio de Suramérica. Este recibió el nombre de virus Río Mamoré y se identificó al Oligoryzomyz microtis como hospedero-reservorio.
Hantavirus en Colombia. Entre 2004 y 2006 se publicaron los primeros estudios serológicos realizados en el Caribe colombiano, en los que se reportó la circulación de hantavirus en humanos (60) y en roedores (61) usando antígenos del 'virus sin nombre' (SNV). Posteriormente, en 2011, se realizó la identificación genética parcial de un probable nuevo hantavirus en roedores del Urabá antioqueño colombiano (62). En 2012 otro estudio demostró la infección en seres humanos con antígenos de hantavirus de las cepas suramericanas Maciel y Araraquara (63). En el departamento de Sucre se realizó un estudio de seroprevalencia de anticuerpos contra hantavirus que evidenció seropositividad a hantavirus en roedores de la subfamilia Murinae. El 8,3 % (12/144) de las capturas fueron seropositivas (64). La expansión del virus hasta Panamá sugiere que la Costa Caribe colombiana, especialmente los departamentos de Córdoba y Sucre, reúnen todas las condiciones que favorecen un eventual evento epizoótico que probablemente podría resultar en un brote de SCPH en humanos, teniendo en cuenta la presencia de grandes extensiones de agroecosistemas, la estación seca marcada y la presencia de roedores reservorios. En ese sentido, existe evidencia indirecta de infección por hantavirus en poblaciones humanas y de roedores reservorios de la Costa Caribe. Sin embargo, en Colombia la hantavirosis no es considerada una enfermedad de notificación obligatoria; y es probable que por desconocimiento y carencia de un diagnóstico diferencial con otras patologías tropicales, esta permanezca actualmente en silencio epidemiológico. En el 2015, a partir de un estudio de fiebres hemorrágicas en el departamento del Meta, se reportó por primera vez en tres pacientes la seroconversión por infección por hantavirus (65). Un análisis filogenético usando como marcador genes precursores de la glicoproteína plantea la tesis de que el virus Necoclí representa una nueva especie en el género hantavirus y que tiene como reservorio a Zygodontomys cherriei (66, 67).
Conclusiones
Las infecciones por hantavirus son un problema de salud pública en muchos países del mundo. En Latinoamérica se han identificado diversos roedores reservorios de cepas patógenas y no patógenas de hantavirus, como también la transmisión de persona a persona del virus Andes. En Colombia se ha descrito la actividad de hantavirus a través de la seroprevalencia en humanos y roedores, por lo cual es importante considerar la búsqueda de casos clínicos compatibles con esta infección que conduzcan al diagnóstico e identificación de los agentes virales autóctonos. Dado que se tiene evidencia serológica en reservorios y casos en humanos en Colombia, se deben tomar medidas de prevención, como evitar el contacto con los roedores y sus excretas, evitar el acceso de los roedores a viviendas, limpieza de zonas contaminadas, educación y recomendaciones a trabajadores y personas que puedan estar expuestas, entre otras. Por otro lado, las políticas de salud demandan un enfoque holístico que integre los enfoques de las investigaciones epidemiológicas y clínicas con estudios ecológicos para generar modelos predictivos que permitan entender aspectos de la bioecología de los reservorios, y esto, a su vez, permita distintas opciones de gestión de la enfermedad.
Referencias
1. Nowak RM. Walker's Mammals of the World. 6th Ed. JHU Press; 1999. [ Links ]
2. Smith MF, Patton JL. Phylogenetic relationships and the radiation of Sigmodontinae rodents in South America: evidence from cytochrome b. Journal of Mammalian Evolution 1999; 6(2): 89-128. [ Links ]
3. Engel SR, Hogan KM, Taylor JF, Davis SK. Molecular systematics and paleobiogeography of the South American Sigmodontinae rodents. Mol Biol Evol 1998; 15(1): 35-49. [ Links ]
4. Fiona A. Red. Rodents. mammals of Central America and Southeast Mexico. 2th ed. Oxford University Press; 2009. p. 186-250. [ Links ]
5. Hjelle B, Torres-Pérez F. Hantaviruses in the Americas and their role as emerging pathogens. Viruses 2010; 2: 2559-2586. [ Links ]
6. Wilson DE, Reeder DM. Mammal species of the world: a taxonomic and geographic reference. 3th ed. Johns Hopkins University Press; 2005. [ Links ]
7. Bennett SN, Gu SH, Kang HJ, Arai S, Yanagihara R. Reconstructing the evolutionary origins and phylogeography of hantaviruses. Trends Microbiol 2014; 22(8): 473-482. [ Links ]
8. Plyusnin A, Sironen T. Evolution of hantaviruses: co-speciation with reservoir hosts for more than 100 MYR. Virus Res 2014; 187: 22-26. [ Links ]
9. Jones KE, Patel NG, Levy MA, Storeygard A, Balk D, Gittleman JL, Daszak P. Global trends in emerging infectious diseases. Nature 2008; 451(7181): 990-993. [ Links ]
10. Khalil H, Hörnfeldt B, Evander M, Magnusson M, Olsson G, Ecke F. Dynamics and drivers of hantavirus prevalence in rodent populations. Vector Borne Zoonotic Dis 2014; 14(8): 537-51. [ Links ]
11. McConnell MS. Hantavirus public health outreach effectiveness in three populations: An Overview of Northwestern New Mexico, Los Santos, Panama, and Region IX, Chile. Viruses 2014; 6(3): 986-1003. [ Links ]
12. Lee JG, Gu SH, Baek LJ et al. Muju Virus, Harbored by Myodes regulus in Korea, Might Represent a Genetic Variant of Puumala Virus, the Prototype Arvicolid Rodent-Borne Hantavirus. Viruses 2014; 6(4): 1701-1714. [ Links ]
13. Ramos C. Los hantavirus causantes de la fiebre hemorrágica con síndrome renal y del síndrome pulmonar. Salud pública de México 2008; 50(4): 334-340. [ Links ]
14. Schmaljohn C, Hjelle B. Hantaviruses: A global disease problem. Emerg Infect Dis 1997; 3(2): 95-104. [ Links ]
15. N′Dilimabaka N, Berthet N, Rougeron V, Mangombi JB, Durand P, Maganga GD, Bouchier C, Schneider BS, Fair J, Renaud F, Leroy EM. Evidence of lymphocytic choriomeningitis Virus (LCMV) in domestic mice in Gabon: Risk of emergence of LCMV encephalitis in Central Africa. J Virol 2015; 89(2): 1456-1460. [ Links ]
16. Reinoso JA, de la Rubia FJ. Spatial spread of the Hantavirus infection. Phys Rev E Stat Nonlin Soft Matter Phys 2015 Mar 91(3): 032-703. [ Links ]
17. Lee HW. Epidemiology and pathogenesis of hemorrhagic fever with renal syndrome. In: Elliott RM, Editors. The Bunyaviridae. New York: Plenum Press; 1996. p. 253-267. [ Links ]
18. Zeier M, Handermann M, Bahr U, Rensch B, Muller S, Kehm R et al. New ecological aspects of hantavirus infection: Change of paradigm and a challenge of prevention a review. Virus Genes 2005; 2:30(2): 157-80. [ Links ]
19. Paul JR, Mc CW. Epidemic hemorrhagic fever attack rates among United Nations troops during the Korean war. Am J Hyg 1958; 68(2): 126-39. [ Links ]
20. Lee HW, Lee PW, Johnson KM. Isolation of the etiologic agent of Korean Hemorrhagic fever. J Infect Dis 1978; 137(3): 298-308. [ Links ]
21. McCormick JB, Sasso DR, Palmer EL, Kiley MP. Morphological identification of the agent of Korean haemorrhagic fever (Hantaan virus) as a member of the Bunyaviridae. Lancet 1982; 1(8275): 765-768. [ Links ]
22. Lee HW, Baek LJ, Johnson KM. Isolation of Hantaan virus, the etiologic agent of Hantaan virus, the etiologic agent of Korean hemorrhagic fever, from wild urban rats. J Infect Dis 1982; 146(5): 638-644. [ Links ]
23. Vapalahti O, Mustonen J, Lundkvist A, Henttonen H, Plyusnin A, Vaheri A. Hantavirus infections in Europe. Lancet Infect Dis 2003; 3(10): 653-661. [ Links ]
24. Settergren B, Juto P, Trollfors B, Wadell G, Norrby SR. Clinical characteristics of nephropathia epidemica in Sweden: prospective study of 74 cases. Rev Infect Dis 1989; 11(6): 921-927. [ Links ]
25. Juto P, Elgh F, Ahlm C, Alexeyev OA, Edlund K, Lundkvist A et al. The first human isolate of Puumala virus in Scandinavia as cultured from phytohemagglutinin stimulated leucocytes. J Med Virol 1997; 53(2): 150-156. [ Links ]
26. Avsic-Zupanc T, Xiao SY, Stojanovic R, Gligic A, van der Groen G, LeDuc JW. Characterization of Dobrava virus: a Hantavirus from Slovenia, Yugoslavia. J Med Virol 1992; 38(2): 132-137. [ Links ]
27. Lee PW, Amyx HL, Gajdusek DC, Yanagihara RT, Goldgaber D, Gibbs CJ, Jr. New hemorrhagic fever with renal syndrome-related virus in rodents in the United States. Lancet 1982; 18:2(8312): 1405. [ Links ]
28. Lee PW, Amyx HL, Yanagihara R, Gajdusek DC, Goldgaber D, Gibbs CJ, Jr. Partial characterization of Prospect Hill virus isolated from meadow voles in the United States. J Infect Dis 1985; 152(4): 826-829. [ Links ]
29. Nichol ST, Spiropoulou CF, Morzunov S, Rollin PE, Ksiazek TG, Feldmann H et al. Genetic identification of a hantavirus associated with an outbreak of acute respiratory illness. Science 1993; 262(5135):914-917. [ Links ]
30. Plyusnin A, Morzunov SP. Virus evolution and genetic diversity of hantaviruses and their rodent hosts. Curr Top Microbiol Immunol 2001; 256: 47-75. [ Links ]
31. Hughes AL, Friedman R. Evolutionary diversification of protein-coding genes of hantaviruses. Mol Biol Evol 2000; 17(10): 1558-1568. [ Links ]
32. Klein SL, Zink MC, Glass GE. Seoul virus infection increases aggressive behaviour in male Norway rats. Anim Behav 2004; 67(3): 421-429. [ Links ]
33. Khalil H, Olsson G, Ecke F, Evander M, Hjertqvist M, Magnusson M, Löfvenius MO, Hörnfeldt B. The Importance of Bank Vole Density and Rainy Winters in Predicting Nephropathia Epidemica Incidence in Northern Sweden. PLoS ONE 2014; 9(11): e111663. doi:10.1371/journal.pone.0111663. [ Links ]
34. Olsson GE, Ahlm C, Elgh F, Verlemyr AC, White N, Juto P et al. Hantavirus antibody occurrence in bank voles (Clethrionomys glareolus) during a vole population cycle. J Wildl Dis 2003; 39(2): 299-305. [ Links ]
35. Lin SL. Parasite manipulation of the proximate mechanisms that mediate social behavior in vertebrates. Physiol Behav 2003; 79(3): 441-449. [ Links ]
36. Nuzum EO, Rossi CA, Stephenson EH, LeDuc JW. Aerosol transmission of Hantaan and related viruses to laboratory rats. Am J Trop Med Hyg 1988; 38(3): 636-640. [ Links ]
37. Easterbrook JD, Kaplan JB, Glass GE, Pletnikov MV, Klein SL. Elevated testosterone and reduced 5-HIAA concentrations are associated with wounding and hantavirus infection in male Norway rats. Horm Behav 2007; 52(4):474-481. [ Links ]
38. Calisher CH, Sweeney W, Mills JN, Beaty BJ. Natural history of Sin Nombre virus in western Colorado. Emerg Infect Dis 1999; 5(1): 126-134. [ Links ]
39. Amman BR, Manangan AP, Flietstra TD, Calisher CH, Carroll DS, Wagoner KD, Mills JN. Association between movement and sin nombre virus (Bunyaviridae: Hantavirus) infection in North American deermice (Peromyscus maniculatus) in Colorado. J Wildl Dis 2013 Jan; 49(1):132-42. [ Links ]
40. Meyer BJ, Schmaljohn CS. Persistent hantavirus infections: characteristics and mechanisms. Trends Microbiol 2000; 8(2): 61- 67. [ Links ]
41. Botten J, Mirowsky K, Kusewitt D, Bharadwaj M, Yee J, Ricci R et al. Experimental infection model for Sin Nombre hantavirus in the deer mouse (Peromyscus maniculatus). Proc Natl Acad Sci USA 2000; 5:97(19): 10578-10583. [ Links ]
42. Klein SL, Bird BH, Glass GE. Sex differences in immune responses and viral shedding following Seoul virus infection in Norway rats. Am J Trop Med Hyg 2001; 65(1): 57-63. [ Links ]
43. Klein S. Host factors mediating sex differences in viral infection. Gend Med 2005; 2(4): 197-207. [ Links ]
44. Kallio ER, Helle H, Koskela E, Mappes T, Vapalahti O. Age-related effects of chronic hantavirus infection on female host fecundity. J Anim Ecol 2015 848(5): 1264-1272. [ Links ]
45. Mills JN, Schmidt K, Ellis BA, Calderon G, Enria DA, Ksiazek TG. A longitudinal study of hantavirus infection in three sympatric reservoir species in agroecosystems on the Argentine Pampa. Vector Borne Zoonotic Dis 2007; 7(2): 229-40. [ Links ]
46. Otteson EW, Riolo J, Rowe JE, Nichol ST, Ksiazek TG, Rollin PE et al. Seroepidemiologic studies of hantavirus infection among wild rodents in California. Emerg Infect Dis 1997; 3(2): 183-190. [ Links ]
47. Teixeira BR, Loureiro N, Strecht L et al. Population Ecology of Hantavirus Rodent Hosts in Southern Brazil. Am J Trop Med Hyg 2014; 91(2):249-257. [ Links ]
48. Klein SL. The effects of hormones on sex differences in infection: from genes to behavior. Neurosci Biobehav Rev 2000; 24(6): 627-638. [ Links ]
49. Easterbrook JD, Zink MC, Klein SL. Regulatory T cells enhance persistence of the zoonotic pathogen Seoul virus in its reservoir host. Proc Natl Acad Sci USA 2007; 25: 104(39): 15502-15507. [ Links ]
50. Schountz T, Prescott J, Cogswell AC, Oko L, Mirowsky-Garcia K, Galvez AP et al. Regulatory T cell-like responses in deer mice persistently infected with Sin Nombre virus. Proc Natl Acad Sci USA 2007; 25:104(39): 15496-15501. [ Links ]
51. Gavrilovskaya IN, Apekina NS, Myasnikov Yu A, Bernshtein AD, Ryltseva EV, Gorbachkova EA et al. Features of circulation of hemorrhagic fever with renal syndrome (HFRS) virus among small mammals in the European U.S.S.R. Arch Virol 1983; 75(4): 313-316. [ Links ]
52. Weber de Melo V, Sheikh Ali H, Freise J et al. Spatiotemporal dynamics of Puumala hantavirus associated with its rodent host, Myodes glareolus. Evol Appl 2015; 8(6):545-559. [ Links ]
53. Douglass RJ, Wilson T, Semmens WJ, Zanto SN, Bond CW, Van Horn RC et al. Longitudinal studies of Sin Nombre virus in deer mouse-dominated ecosystems of Montana. Am J Trop Med Hyg 2001; 65(1): 33-41. [ Links ]
54. Boone JD, McGwire KC, Otteson EW, DeBaca RS, Kuhn EA, Villard P et al. Remote sensing and geographic information systems: charting Sin Nombre virus infections in deer mice. Emerg Infect Dis 2000; 6(3): 248-258. [ Links ]
55. Yan L, Fang LQ, Huang HG, Zhang LQ, Feng D, Zhao WJ et al. Landscape elements and Hantaan virus-related hemorrhagic fever with renal syndrome, People's Republic of China. Emerg Infect Dis 2007; 13(9): 1301-1306. [ Links ]
56. Lazaro ME, Cantoni GE, Calanni LM, Resa AJ, Herrero ER, Iacono MA et al. Clusters of hantavirus infection, southern Argentina. Emerg Infect Dis 2007; 13(1): 104-110. [ Links ]
57. Padula PJ, Edelstein A, Miguel SD, Lopez NM, Rossi CM, Rabinovich RD. Hantavirus pulmonary syndrome outbreak in Argentina: molecular evidence for person-to-person transmission of Andes virus. Virology 1998; 15:241(2): 323-330. [ Links ]
58. Martinez VP, Bellomo C, San Juan J, Pinna D, Forlenza R, Elder M et al. Person-to-person transmission of Andes virus. Emerg Infect Dis 2005; 11(12): 1848-1853. [ Links ]
59. Klempa B, Fichet-Calvet E, Lecompte E, Auste B, Aniskin V, Meisel H et al. Novel hantavirus sequences in Shrew, Guinea. Emerg Infect Dis 2007; 13(3): 520-522. [ Links ]
60. Máttar S, Parra M. Serologic evidence of hantavirus infection in humans, Colombia. Emerg Infect Dis 2004; 10(12): 2263-2264. [ Links ]
61. Alemán A, Iguarán H, Puerta H, Cantillo C, Mills J, Ariz W et al. First serological evidence of hantavirus infection in rodents in Colombia. Rev. Salud Pública 2006; 8(1): 1-12. [ Links ]
62. Londoño AF, Díaz FJ, Agudelo FP, Levis S, Rodas JD. Genetic evidence of hantavirus infections in wild rodents from northwestern Colombia. Vector Borne Zoonotic Dis 2011; 11(6): 701-708. [ Links ]
63. Guzmán C, Mattar S, Levis S, Pini N, Figueiredo LT, Mills J, Salazar-Bravo J. Prevalence of antibody to hantaviruses in humans and rodents in the Caribbean region of Colombia determined using Araraquara and Maciel virus antigens. Mem Inst Oswaldo Cruz 2013; 108: 167-171. [ Links ]
64. Blanco P, Arroyo S, Corrales H, Pérez J, Álvarez L, Castellar A. Evidencia serológica de infección por hantavirus (Bunyaviridae: Hantavirus) en roedores del departamento de Sucre, Colombia. Rev. Salud Pública 2012; 14 (5): 755-764. [ Links ]
65. Sánchez L, Mattar S, Rodríguez I, Rodríguez D, Tiquet V. First serological evidence of Hantavirus infection. In humans from the Orinoquia region of Colombia. Zoo Pub Heath 2015 (In press). [ Links ]
66. Montoya RC, Díaz FJ, Rodas JD. Recent evidence of Hantavirus circulation in the American Tropic. Viruses 2014; 6:1274-1293. [ Links ]
67. Montoya RC, Cajimat MNB, Louise Milazzo ML, Diaz FJ, Rodas JD, Valbuena G, Fulhors CF. Phylogenetic Relationship of Necoclí virus to other South American Hantaviruses (Bunyaviridae: Hantavirus). Vector-Borne and Zoonotic Diseases 2015; 15(7):438-445. [ Links ]

