











 Spanish (pdf)
Spanish (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mail Cited by SciELO
Cited by SciELO  Cited by Google
Cited by Google  Similars in
SciELO
Similars in
SciELO  Similars in Google
Similars in Google 
 Permalink
Permalink
Introducción
Prochilodus mariae (bocachico llanero o coporo), es una especie de la cuenca del Orinoco, tiene gran importancia tanto ecológica como comercial en ecosistemas acuáticos, siendo la segunda especie de mayor importancia con un aporte de 153.4 toneladas/año (MADR y CCI, 2010). Se han realizado estudios en pesquerías, manejo de primera alimentación en cultivo, pero aún existen vacíos de información en el conocimiento por la preferencia del nivel trófico durante los primeros días de vida. Arias-Gallo et al., (2010) determinaron que es común en larvas pelágicas, al iniciar la alimentación exógena posean, una mandíbula funcional y un desarrollo del tracto digestivo, que le permita a la larva reaccionar al estímulo lumínico, detectar presas perifíticas, zooplanctónicas y predadores potenciales en la columna de agua y comenzar a alimentarse cuando las reservas del saco vitelino sean absorbidas. López et al., (2005), determinaron que estructuras como la boca y el intestino se desarrollan durante la vida larval, aun antes de reabsorber por completo el saco vitelino, situación que permite el inicio de la alimentación exógena. Sánchez et al., (2003), la definen como especie detritívora o sedibentófaga, con boca protráctil y labios gruesos que le ayudan a succionar sedimentos, además posee branquiespinas papilosas, estómago reducido especializado con molleja de pared muscularizada. Esto último les permite triturar mediante granos de arena, el material orgánico, además presenta un intestino muy largo y numerosos ciegos pilóricos que incrementan el área de absorción. Estas características les permiten optimizar la deglución y absorción de materiales detríticos y elementos orgánicos poco particulados. Atencio-García et al., (2003), realizaron un estudio en Prochilodus magdalenae cuyos resultados sugieren que nauplios de artemia o zooplancton son presas adecuadas para el manejo de la primera alimentación. Beltrán et al., (2001). Consideran que el coporo es una especie importante ecológicamente porque se ubica en la base de la red trófica ya que es detritívora, cumpliendo un rol fundamental en el ciclo de nutrientes, debido a que toma la materia orgánica y los minerales de los sedimentos y los hace disponibles a las otras especies ícticas.
Donoso et al., (1997), en trabajo realizado con P. magdalenae, concluyeron que a medida que aumenta el tamaño de la pos larva de bocachico, disminuye el tamaño de la partícula ingerida, tendiendo una preferencia por partículas finas, que más adelante le determinaran la condición de consumidor de detrito.
Bustamante et al., (1997), recomiendan proporcionar alimento vivo a las larvas a partir del momento que inician nado horizontal, para evitar mortalidades al momento de la siembra en los estanques en tierra. P. mariae, en la primera semana consumen rotíferos, luego muestran preferencia por los cladóceros, cuando no tienen una fuente de zooplancton suplen sus necesidades con algas de diversos tipos.
El factor limitante para el desarrollo de su piscicultura se ha presentado en la producción de alevinos, debido a la alta tasa de mortalidad en la primera fase de crecimiento, problema que se deriva de la falta de conocimiento sobre la alimentación de las larvas y las poslarvas en condiciones naturales que puedan dar los soportes científicos y técnicos para la cría de estas en confinamiento (Arias, 1995). En este estudio, se aportan bases que permiten avanzar en el desarrollo de la piscicultura de P. mariae, evaluando las preferencias alimenticias en etapas iniciales.
Materiales y métodos
Ubicación del experimento
Los experimentos se desarrollaron en la Estación Piscícola del Instituto de Acuicultura de los Llanos, en tanto que los estudios de los organismos en el Laboratorio de Biología de la Facultad de Ciencias Básicas e Ingeniería, de la Universidad de los Llanos en Villavicencio, Meta, Colombia.
Material biológico
Larvas obtenidas mediante reproducción inducida fueron sembradas a las 72 horas pos-eclosión (HPE), cuando presentaban apertura oral y anal, natación horizontal y un remanente del 20% del saco vitelino, en dos estanques de tierra de 250 m2 preparados previamente simulando condiciones naturales como sigue; los estanques secos durante tres semanas fueron abonados uniformemente con bovinaza seca a razón de 10 g/m2 y 2 g/m2 de heno comercial Pennisetum clandestinum, y luego llenados lentamente durante 7 días al cabo de los cuales se sembraron 20 larvas / m2.
Desde el día de la siembra, cada dos días hasta por 30 días, a las 8:00 h, fueron muestreadas las aguas y los organismos. Los parámetros de calidad de agua; (temperatura, pH, registrados con el pHmetro portátil Hanna instruments (pH/ORP/Temp). Oxígeno disuelto, con el oxímetro portátil HI98196 Hanna instruments. Conductividad con un conductímetro portátil (0,0 μS/cm) Hanna. Dureza, (Test kit dureza total Hanna, rango 0,0-53 mg CaCO3). Alcalinidad y acidez HI3813. Fueron registrados según Boyd (1996). Para el muestreo de plancton y fitoplancton se aplicó la metodología de Gaviño, et al., (1982) citado por Sánchez et al., 2013, fue colectado con red de 60 μm de ojo de malla, hasta completar 10 L, luego este fue concentrado en un tamiz de 15 μm hasta un volumen de 10 mL y fijado, inmediatamente, con formol bufferado al 4%. Para evaluar el perifiton se aplicó la metodología propuesta por Roldán (2008), se tomó una superficie foliar de 16±1 cm2, de heno colectado en cada caso, fueron lavados cuidadosamente y desprendidos los organismos y concentrados en 10 mL y fijados con 0.4 ml de formol bufferado del 4% más 0.4 ml de solución de lugol. Nueve poslarvas/estanque/muestreo, fueron colectados y fijados en formol bufferado al 4 %. Los organismos de cada muestreo descrito fueron contados con la ayuda de una cámara de microfotografía NIKON Digital, SIGHT DS-U1, Japón, acoplada a un microscopio de contraste de fase (NIKON, Japón). Para la identificación y determinación de los microorganismos se siguió a Heinz y Dieter (1987). La abundancia del perifiton se determinó teniendo en cuenta el número de individuos/área. Para la abundancia del plancton pelágico se tomaron tres sub-muestras de 0.05 ml del concentrado, las que fueron colocadas en cámara de Neubauer y luego observadas al microscopio óptico para registrar un inventario de todos los organismos presentes. Para las muestras de bentos se analizaron aplicando la metodología descrita por Roldán (2008). En poslarvas con tubo digestivo no diferenciado se analizó el contenido total y en los individuos más desarrollados se estudió solo el contenido estomacal. Para el análisis de los contenidos intestinales y estomacales se retiró el tubo digestivo de cada individuo y luego se raspó y se colocó el contenido en una cámara de Neubauer a la que se le agregó 0.05 mL de agua destilada. Las observaciones bajo el microscopio siguieron los procedimientos descritos por (Hyslop, 1980; Rossi, 1992; Atencio et al., 2003).
Los registros fueron agrupados por tallas de individuos, por fechas y por desarrollo aparente del tubo digestivo v/s ítems alimenticios, y con ello se elaboraron tablas de análisis con índice de frecuencia (IF) de los contenidos y la oferta de los microorganismos en el estanque por origen (perifiton, plancton, bentos), en porcentaje de la oferta.
El Índice de Frecuencia de los contenidos estomacales se calculó, según Mar-Silva et al., (2014), aplicando la fórmula: IF = n/Ne
Resultados
En la Tabla 1, se observan los parámetros físico-químicos evaluados en los estanques durante el período de muestreo. Los resultados presentaron muy pocas variaciones, T°: 26.5±0.5, pH: 6.3±0.5, OD: 6.2±0.3mg/l, alcalinidad: 11.5±3mg/l, dureza: 16±8mg/l CaCO3 y Conductividad: 15±4 μS/cm.
Tabla 1 Parámetros físico-químicos del agua en los dos estanques. T°, temperatura OD, oxígeno disuelto.

Como se puede apreciar en la Tabla 2, las poslarvas con intervalo de 6 - 8.99 mm, de longitud total (LT) mostraron una tendencia por el perifiton, en los contenidos estomacales (CE), presentaron un 15.9% de algas clorofíceas. Un 66.7% de las poslarvas muestreadas presentaron estómagos vacíos. En la Tabla 3, los contenidos estomacales de las poslarvas con intervalos 9-11.99; 12-14.99 mm de LT, presentan en los CE, microcrustáceos (cladóceros y copépodos) en porcentajes de 50.7 y 72.0% ofertados, en su gran mayoría, por el plancton pelágico. El bentos aporta un porcentaje secundario como alimento pero en los CE, no se observó presencia de este.

Tabla 3 Abundancia de los diferentes ítems alimenticios en los contenidos estomacales (C.E) de las poslarvas de P. mariae, de 6,0 - 14,9 (mm) de longitud total y el aporte de los diferentes niveles tróficos (perifiton, plancton, bentos).
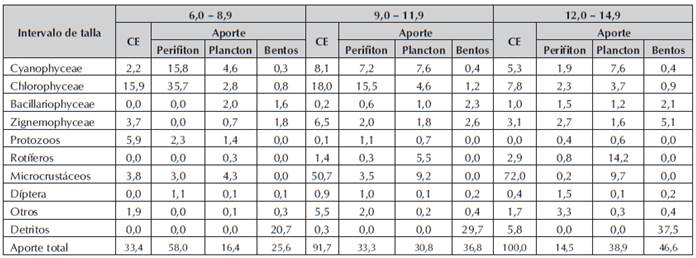
En los intervalos de 15-17.99; 18-20.99; 21-23.99 mm de LT (Tabla 4), se observó un incremento en el consumo de alimento de tipo secundario por algas Zignemophyceae, ofertadas por el bentos. Las poslarvas presentaron una tendencia a consumir detritos incrementándose a medida que aumenta la longitud total de las mismas.
Tabla 4 Abundancia de los diferentes ítems alimenticios en los contenidos estomacales (C.E) de las poslarvas de P. mariae, de 15,0 - 23,9 (mm) de longitud total y el aporte de los diferentes niveles tróficos (perifiton, plancton, bentos).

En las tablas 5 y 6, se observan las poslarvas con intervalos de 24-26.99; 27-29.99, 30-32.99; 33-35.99; 36-38.99 y 39-41.99 mm de LT, las cuales mostraron un mayor consumo de algas Zignemophyceae y de detritos ofertados en su mayor parte por el bentos.
Tabla 5 Abundancia de los diferentes ítems alimenticios en los contenidos estomacales (C.E) de las poslarvas de P. mariae, de 24,0 - 32,9 (mm) de longitud total y el aporte de los diferentes niveles tróficos (perifiton, plancton, bentos).

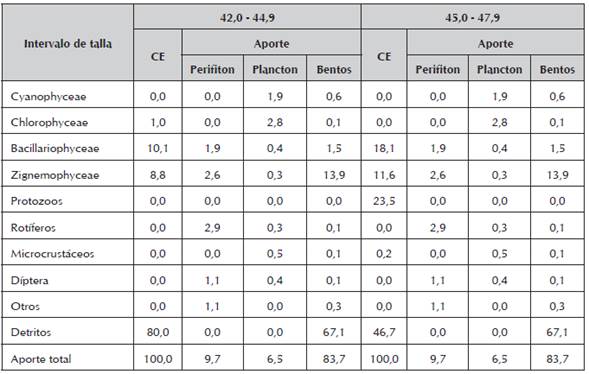
Los alevinos con intervalo de 42-44.99 y 45-47.99 mm de LT, presentaron un consumo de algas Bacillariophyceae de tipo accidental (10 y 18%) y preferencia por los detritos. Los dos ítems fueron ofertados por el bentos (Tabla 6), el cual representó el 83,7% de la ingesta alimenticia.

Discusión
En el presente trabajo se estudiaron las preferencias alimenticias por las poslarvas y el inicio de alevinaje de P. mariae, en estanques en tierra que fueron abonados orgánicamente con bovinaza y heno, condiciones simuladas a las presentadas en zonas de rebalse y/o llanuras de inundación, permitiendo el desarrollo de comunidades perifíticas, planctónicas y de bentos. Según Donato (1987), reporta que en el ambiente natural, luego de la época de sequía, se inicia la época anegada, donde las comunidades de perifiton cambian en su composición y estructura. Las especies dominantes del perifiton están rodeadas de copiosas envolturas de mucílago, como se pudo observar en Microcystis, Sphaerocystis, Eremosphaera, encontradas en este trabajo.
En este estudio, las poslarvas con intervalo de 6-8.99 mm de LT, comenzaron su alimentación exógena. Rossi (1992) observó que Prochilodus lineatus inició alimentación exógena a partir de 7.5 mmm de longitud total. Un 66.67% presentaron estómagos vacíos (Tabla 3), probablemente porque conservaban un remanente de saco vitelino. Las poslarvas consumen perifiton debido a la poca agilidad y experticia para capturar el plancton pelágico (López et al., 2005). Los detritos ofertados por el bentos representan el 20.7% pero en los contenidos estomacales todavía no hay presencia de ellos, debido a que presentan un tubo digestivo muy incipiente, sin un verdadero estómago muscular (molleja) que le ayude a triturar estos detritos (Sánchez et al., 2003).
Las poslarvas de 9-11.99; 12-14.99; 15-17.99 mm de LT, son muy activas y nadan libremente, presentaron preferencia por el consumo de microcrustáceos, preferiblemente cladóceros y copépodos; Rossi (1992), afirma que los cladóceros, constituyen una parte importante de la biomasa de comunidades en el plancton. Se destacan por su transferencia de materia y energía desde los microproductores primarios y las fuentes de detritos y bacterias hacia los niveles de los consumidores superiores. Rossi (1992) y Paggi (2005), destacan que los cladóceros son alimento importante en poslarvas de especies que en estado ocupen otros nichos tróficos, como es el caso de P. mariae, es importante resaltar que las poslarvas en estas tallas mostraron una tendencia por el plancton pelágico, como se puede observar en la Figura 1.

Figura 1 Aporte de contenidos estomacales según los intervalos de talla de las poslarvas de P. mariae
Las poslarvas con intervalo de 18-20.99; 21-23.99; 24-26.99; 27-29.99 mm de LT (Tabla 4), presentan una tendencia a consumir algas Zignemophyceae (44.9%), como alimento secundario ofertadas por el bentos, siendo los géneros Closterium (55 %), Cosmarium (44 %), Staurastrum (0.5 %), Spirogyra (0.2 %) y Bambusina (0.1 %) los más consumidos, debido a lo expuesto anteriormente. Los individuos de estos intervalos de LT, inician un cambio por la oferta alimenticia mostrando una tendencia por el consumo de las comunidades del bentos.
Las poslarvas con intervalo de 30-32.99; 33-35.99; 36-38.99 y 39-41.99 mm de LT (Tabla 5 y 6), mostraron preferencia por el consumo de bentos, notándose a su vez, incremento en la ingesta de Zignemophyceae y Bacillariophyceae con los géneros Pinnularia (43.2%), Navícula (20.7%), Frustulia (32.8%) y Tabellaria (3.2%). En la Tabla 7, los individuos con tallas entre 42-44.99 y 45-47.99 mm de LT, con 30 días de edad (alevinos) mostraron características morfológicas y anatómicas igual que los adultos, con un nivel trófico detritívoro, observándose en sus CE una preferencia por bacterias, hongos y materia orgánica particulada. Estos resultados coinciden con lo reportado por Sánchez et al., (2013), quienes afirman que los microorganismos, principalmente bacterias, son el alimento de apoyo para detritívoros de partículas finas, siendo una alta calidad de recurso alimenticio.
Tabla 7 Abundancia de los diferentes ítems alimenticios en los Contenidos Estomacales de las poslarvas de P. mariae, de 42,00 - 47,99 (mm) de longitud total y el aporte de los diferentes niveles tróficos (perifiton, plancton, bentos).
En el presente estudio se pudo determinar que existe una preferencia alimentaria por P. mariae, al cultivarlo en estanques en tierra, donde se definen tres cambios en los niveles tróficos, iniciando con una tendencia por el consumo de perifiton (Clorofíceas y Protozoos), una segunda fase con preferencia por microcrustáceos (cladóceros y copépodos) y por último individuos detritívoros con preferencia por el consumo de materia orgánica particulada (bacterias y hongos).

