











 English (pdf)
English (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mail Cited by SciELO
Cited by SciELO  Cited by Google
Cited by Google  Similars in
SciELO
Similars in
SciELO  Similars in Google
Similars in Google 
 Permalink
Permalink
In humans, early-life rearing conditions characterised by nurturing, absent or neglectful parental care can have deleterious effects on neurobehavioural development (Nelson et al., 2023). Experimental research with animal models has facilitated a better understanding of the long-term physiological and behavioural effects of children’s exposure to early adverse experiences (Nishi, 2020; Smith & Pollak, 2020). Maternal separation (MS) is a commonly used animal model to assess the effects of early-life stress, such as child neglect (Sun et al., 2021). In general, MS consists of separating the pups from their dams during the first weeks of postnatal life, a stage in which the pups depend on maternal care to survive (Nishi, 2020; Halladay & Herron, 2023). Similar to the consequences reported in humans, rodent pups separated from their dams exhibit long-term physiological and behavioural alterations, such as changes in the functioning of the hypothalamic-pituitary-adrenal axis and the stress response (Eskandari et al., 2023).
Communal nesting is a paradigm used in mice to study the long-term effects of early social enrichment, consisting of a single nest in which a group of dams share caring behaviours from birth to weaning (Branchi & Alleva, 2006). The maternal care exhibited by dams in the communal nesting condition is greater than in the standard single-female-per-nest condition (D’Andrea et al., 2010). Early-rearing of mice in communal nests made them resistant to exhibit depression-related behaviours (D’Andrea et al., 2010). The communal nesting paradigm as a model of early social enrichment has also been described in Norway rats (Martinez et al., 2015), Sprague-Dawley rats (Connors et al., 2015) and recently in Wistar rats (Bratzu et al., 2023).
The elevated plus-maze (EPM) has been an extensively used test to assess anxiety-related behaviours in rodents and is based on rats’ natural aversion to open spaces (Kraeuter et al., 2019). In EPM, conventional spatiotemporal measures such as time spent in the open arms and entries into closed arms have been interpreted as measures related to anxiety and locomotion, respectively (Carobrez & Bertoglio, 2005). Additionally, other behaviours such as increased stretch attend postures and vertical exploration are indicative of anxiogenic-like effect (Holly et al., 2016; Kuniishi et al., 2017). It was shown that the reduction of time in the central area of the EPM correlated with an impulsive phenotype during an operant task. Thus, the time in the central area could be considered as a parameter of motor impulsivity in rats (Rico et al., 2016).
Maternally separated adult rats display an increase in anxiety-related behaviours when exposed to the EPM (Wang et al., 2020). In the same way, adult mice reared in communal nesting showed higher levels of anxiety-like behaviour in the EPM, compared to control mice (Branchi & Alleva, 2006). It has been described in rats that early exposure to social enrichment by communal nesting results in a low-anxiety behavioural phenotype assessed in an open field, either during adolescence (Bratzu et al., 2023) or in adulthood (Martinez et al., 2015). However, a high-anxiety phenotype due to early social enrichment has also been reported in adolescents exposed to EPM (Connors et al., 2015).
To our knowledge, no study has compared two opposite early-rearing conditions on the behavioural profile of rats at the onset of adolescence. Thus, we evaluated the effects of maternal separation and communal nesting on emotional response of adolescent male and female rats exposed to the EPM test. We hypothesise that early rearing with MS might result in an increased anxiety phenotype, whereas SE by community nesting might have an opposite effect at the onset of adolescence.
Method
Subjects
Six Wistar male and twelve nulliparous female rats, obtained from the National Institute of Health (Bogotá, Colombia) were mated overnight and separated the following morning. The pregnant rats were housed in polycarbonate cages (42 × 20 × 20 cm) which contained dust-free wood shaving bedding and were kept in an animal room under a 12h light/dark cycle (lights turned on at 07:00) with water and standard rodent pellets available ad libitum. Room temperature was maintained at 23°C with 55% relative humidity. All experimental procedures were performed following the United States National Institute of Health Guide for the Care and Use of Laboratory Animals and were approved by our Institutional Animal Care and Use Committee (CICUAL-KL; COM27-21-07-2015).
Early-rearing procedures
Ten females confirmed to be pregnant were randomly distributed into three groups according to early-life rearing conditions: Standard Housing (SH; n = 2), Maternal Separation (MS; n = 2), and Social Enrichment (SE; n = 6). The day of birth was designated as postnatal day (P) 0. On P1, litters were culled to eight pups (4 males and 4 females). Offspring were weaned at P23 and housed with same-sex littermates.
Standard housing (SH). The two pregnant primiparous females were individually housed in standard cages and kept with their offspring until weaning. Only procedures related to cage cleaning and weight recording were performed.
Maternal separation (MS). The two pregnant primiparous females were individually housed in standard cages and kept with their offspring until weaning. Pups were separated from their dams for 180 minutes in the morning (9:00 to 12:00h) from P2 to P14. During separation, the litters were placed individually in a separate room on a heating pad. After 180 minutes, the litters were returned to their dam’s cage.
Social enrichment (SE). A communal nesting procedure for rats, similar to that described by Connors et al. (2015), was used as social enrichment. The six pregnant primiparous females were pair-housed in standard cages. Of the six females, only four that gave birth synchronously within a 24-hour period were considered. Thus, each of the two cages consisted of a pair of dams and eight pups (4 males and 4 females). Only procedures related to cage cleaning and weight recording were performed.
During P14-P32, the body weight was measured with a scale once every three days. In order to minimise the impact of the variations in initial body weight within the groups, the percentage of weight gain with respect to P14 was calculated.
Behavioural procedures
In order to obtain locomotor, vertical exploration, anxiety and impulsive-related measures, an elevated plusmaze (EPM) was used. The instrument had a centre zone (10 x 10-cm), two open arms (50 x 10-cm) with a side border (0.5 cm), and two closed-arms (50 x 10-cm) with 40-cm high black Plexiglas walls. The floor was made of black Plexiglas and the instrument was elevated 50cm from the floor. The lighting at the centre of the maze was adjusted to 30 lux.
On P32, when the animals reached the onset of adolescence (Arellano et al., 2024), three rats from each same-sex littermate cage were randomly selected for behavioural testing. Thirty-six animals (SH, MS and SE; n = 12) divided by sex (n = 6) were exposed to the EPM. Rats were transported from their living room to the experimental room using a transport cage. Each animal was carefully placed in the central area of the EPM facing one closed arm, allowing it to explore the instrument for 5 minutes. At the end of each session, the instrument was cleaned using 10% alcohol and dried with paper towels.
All sessions were recorded by a video camera placed above the EPM and connected to a digital video recorder in an adjacent room. Videos were subsequently analysed by a highly trained observer, being aware of the analysed factors. Behavioural measures were scored with the ethological scoring free software X-PloRat (Tejada et al., 2018). The scored behaviours included the percentage of time spent in the open arms and the central area, the number of entries in the open and closed arms, the frequency of vertical exploration and the time spent in stretch-attend postures (SAP). An entry into an arm was scored after all four paws of the rat entered it. Vertical activity was recorded whenever the rat leaned against the wall with its forelegs in a high position (Kuniishi et al., 2017). SAP were scored when the rat lowers its back, elongates its body, and is either standing still or moving forward very slowly (Holly et al., 2016).
Statistical analysis
Levene’s test was used to verify data equality of variances and when equality was confirmed the statistical study was performed by way of analysis of variance (ANOVA) complemented by a Tukey test. Data were shown as the mean ± SEM. The percentage of weight gain for male and female rats was analysed using twoway repeated measures ANOVA (Early-rearing x weight days). The EPM data were analysed using two-way ANOVA (Early-rearing x Sex). The level of significance was set at p < .05.
Results
Two-way repeated measures ANOVA showed significant interactions between weight days and early-rearing condition in male (F(10,75) = 3.644; P < .001) and female rats (F(10,75) = 2.734; P < .01). Post hoc tests showed that during P29 and 32, males and females significantly increased body weight compared to P17, 20, 23 and 26 (Figure 1A and 1B). Post hoc tests also showed that during P32, maternally separated females had a significant increase in body weight gain compared to socially enriched ones (Figure 1B).
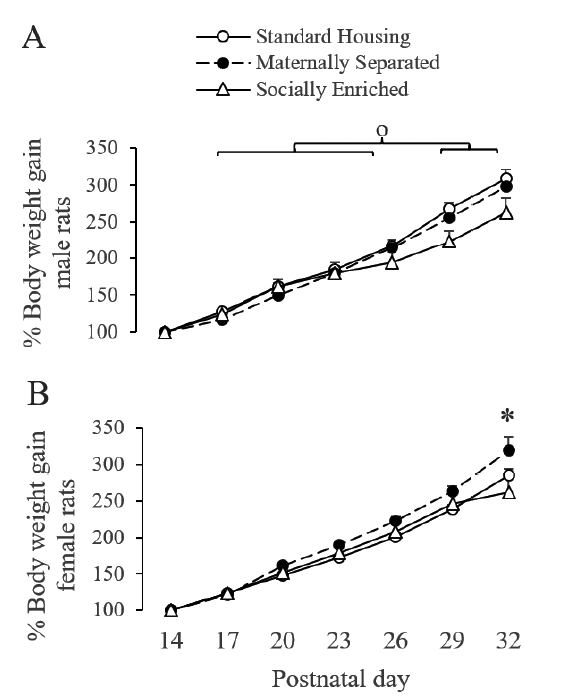
Figure 1 Effect of maternal separation and social enrichment on body weight gain of male and female adolescent rats o During P29 and 32, males and females significantly increased body weight compared to P17, 20, 23 and 26 (p < .05). * During P32, maternally separated females had a significant increase in body weight gain compared to socially enriched female rats (p < .05).
Results from the elevated plus-maze test are summarised in Figures 2 and 3. Two-way ANOVA showed an early-rearing condition effect on the percentage of time spent in the open arms (F(2,30) = 4.562; P < .05) and in the central area (F(2,30) = 8.351; P < .005); the time spent in stretch-attend postures (F(2,30) = 3.446; P < .05), the frequency of entries into the open (F(2,30) = 11.281; P < .001) and into the closed arms (F(2,30) = 7.330; P < .005). Post hoc tests showed that, compared to the standard housing group, maternally separated rats reduced the time spent in the open arms (Figure 2A), the entries into the open arms (Figure 2B) and the time spent in the central area (Figure 3B). Post hoc tests also showed that socially enriched rats increased the time spent in SAP (Figure 2C) and the entries into the closed arms (Figure 2D), compared to the control standard housing group.
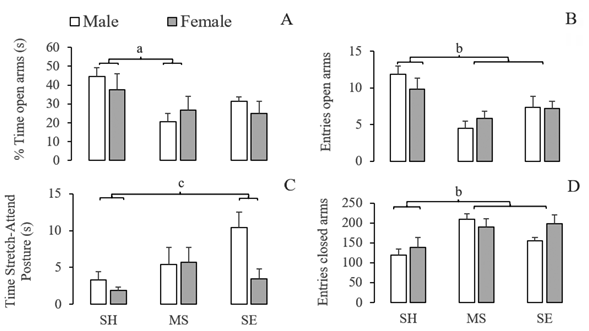
Figure 2 Effect of maternal separation and social enrichment on emotional response of male and female rats in the elevated plus-maze SH: Standard Housing. MS: Maternal separation. SE: Social Enrichment. a Standard housing different from maternal separation (p < .05). b Standard housing different from maternal separation and social enrichment (p < .005). c Standard housing different from social enrichment (p < .05).
Two-way ANOVA also showed a sex effect on the frequency of vertical exploration (F(1,30) = 6.543; P < .05).
Post hoc tests showed that female rats increased vertical exploration compared to male ones. Finally, twoway ANOVA showed a significant interaction between early-rearing conditions and sex on the percentage of time spent in the central area (F(2,30) = 4.802; P < .05) and vertical exploration (F(2,30) = 4.681; P < .05). Post hoc tests showed that socially enriched female rats increased the frequency of vertical exploration (Figure 3A) and reduced the time spent in the central area (Figure 3B) compared to socially enriched male rats and to control female rats.
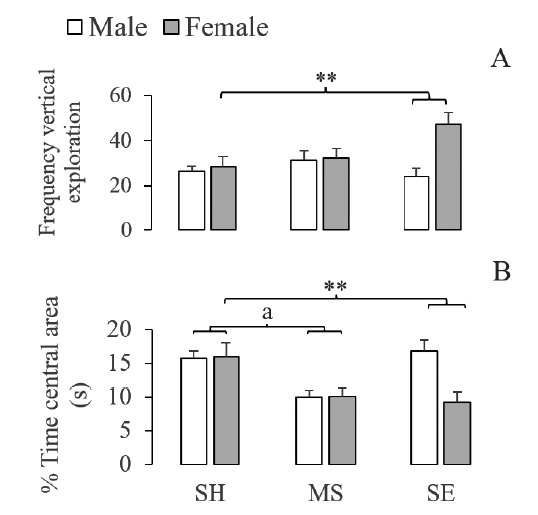
Figure 3 Effect of maternal separation and social enrichment on locomotion (A) and impulsive-related measures (B) in the elevated plus-maze SH: Standard Housing. MS: Maternal separation. SE: Social Enrichment. a Standard housing different from maternal separation (p < .005). ** Socially enriched female different from socially enriched male rats and standard housing female rats (p < .05).
Discussion
This study aimed to determine the effect of two opposing early-life rearing conditions, maternal separation and social enrichment, on the emotional response of male and female rats evaluated at the onset of adolescence in the EPM. Consistent with our initial hypothesis, the results showed that maternal separation resulted in an increased anxiety phenotype in early adolescence. However, social enrichment by communal nesting did not have the expected opposite effect of reducing anxiety-related behaviours. In fact, the behavioural profile of adolescents raised with maternal separation and social enrichment was similar. The results also showed that early rearing conditions had sex-dependent effects on both emotional measures and body weight.
Compared with animals that had been in standard housing conditions, adolescent rats that had been reared with maternal separation and early social enrichment reduced open-arm exploration while increasing entries into the closed arms of the EPM. Results suggest that both early rearing conditions may increase anxiety-related behaviours and locomotion in the EPM. Our results are in agreement with data from the review presented by Wang et al (2020) showing that, in 24 studies, maternal separation was associated with increased defensive behaviour in rats evaluated on the EPM.
Likewise, the increase in anxiety-related behaviours due to early social enrichment that we observed is in agreement with findings reported for mice by Branchi and Alleva (2006) as well as for adolescent Sprague-Dawley rats (Connors et al., 2015). However, our observations do not agree with results found in adolescent Wistar rats (Bratzu et al., 2023) nor in Norway adulthood rats (Martinez et al., 2015) reared with early social enrichment and that showed a low-anxiety behavioural phenotype when exposed to an open field. The discrepancy between the reported results could be partly explained by the behavioural test used to obtain anxiety-related measures (EPM; open field) or by the rat strain used (Wistar; Sprague-Dawley; Norway). However, it is likely that variables related to social enrichment protocol such as the number of dams per communal nest may have influenced the anxiety phenotypes found. While in the studies of Bratzu et al. (2023) and Martinez et al. (2015) the communal nest was formed by three dams, in our protocol and in Connors et al. (2015) just two dams per nest were used. To our knowledge, there is no evidence demonstrating a relationship between the number of females in a communal nest and changes in the anxiety phenotypes of their offspring in rats. The effect of the number of dams in communal nests on the emotional response of their offspring at other stages of development is an issue that needs to be examined in further studies.
Different from standard and maternal separation conditions, sex-differences in adolescents that had been reared with early social enrichment were observed. Under communal nesting conditions, females but not males increased vertical activity and reduced the time in the central area of the EPM. Although not significantly, males but not females increased stretch-attend postures. The results suggest that early social enrichment may have sex-dependent effects on anxiety-related measures, increasing vertical risk assessment in females and horizontal risk assessment in males. Different from males, female rats reared in a communal nest also showed a significant increase in vertical exploration when exposed to an open field during adolescence (Bratzu, 2023). However, communal nesting did not result in that behavioural phenotype assessed in the EPM (Connors et al., 2015). Sex-differences in rats that had been maternally separated were also observed. On P32, females that had been reared with maternal separation showed significant body weight gain compared to socially enriched female rats. Consistent with our findings, other studies also showed that maternal separation increased body weight gain (Ruiz et al., 2018; Machado et al., 2022). However, we did not find weight gain in community nest-reared animals as reported in mice (Heiderstadt & Blizard, 2011).
In conclusion, the results suggest that two early rearing conditions may have similar effects on the emotional response of adolescent rats. Seemingly opposite, maternal separation and communal nesting resulted in increased anxiety and locomotion at the onset of adolescence. In the social enrichment model, the number of dams in the communal nest affects emotional response. Apparently, three but not two females per nest could reduce the anxiety response of adolescent rats in the EPM.
Some methodological aspects limited our study and could be addressed in subsequent research. The behaviour of females towards their offspring in the condition of maternal separation and communal nesting should have been recorded in order to better explain the increased anxiety phenotypes we observed during adolescence. Likewise, other behavioural tests such as open field or social interaction could have been used to obtain a more robust behavioural phenotype derived from the rearing conditions evaluated.

