











 Spanish (pdf)
Spanish (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mail Cited by SciELO
Cited by SciELO  Cited by Google
Cited by Google  Similars in
SciELO
Similars in
SciELO  Similars in Google
Similars in Google 
 Permalink
Permalink
INTRODUCCIÓN
El término "Wagyu" significa ganado japonés, por lo que este término incluye a las cuatro razas de ganado bovino de origen japonés, (Negro japonés [BJ], Café japonés, Polled japonés y Shorthorn japonés) 1. La carne Wagyu es reconocida mundialmente por el alto contenido de grasa intramuscular (IMF), sin embargo, de las cuatro razas, solo el BJ puede producir carne al nivel de marmoleo con el que es reconocida esta carne. Debido a esto, en Japón, BJ ocupa mas del 90% de la población total de ganado y es la única que se cría a gran escala en países como Australia, España y Estados Unidos 2. El término Wagyu fuera de Japón habitualmente se utiliza para referirse a la carne del ganado BJ.
La IMF o marmoleo está estrechamente relacionada a las percepciones sensoriales del consumidor, por lo que es utilizada como un indicador visual de calidad sensorial 3, y el incremento de IMF en la carne se ha asociado a con una mayor palatabilidad 4,5. Igualmente, la composici ón de ácidos grasos (FA) en la IMF influye sobre características sensoriales y nutricionales. En razas específicas, la carne bovina marmoleada no solo tiene una excelente calidad sensorial, sino que también contiene una gran cantidad de FA beneficos para la salud 6.
Generalmente, los ácidos grasos saturados (SFA) han sido relacionados con efectos negativos en la salud de los consumidores y sabor de la carne 7,8; por el contrario, los ácidos grasos monoinsaturados (MUFA), se han asociado a efectos positivos sobre la salud 6 y a las caracteristicas sensoriales, y los ácidos grasos poliinsaturados (PUFA) se han asociado a diversos efectos positivos en la salud. Se ha sugerido que los consumidores prefieren y demandan carne con mayores cantidades de MUFA y menores de SFA 8. Debido a lo anterior, se busca mejorar la proporción MUFA/SFA. Algunos estudios han indicado que la carne Wagyu, contiene mayor concentración de MUFA y menor SFA respecto a Angus, Charolais, Holstein 9,10.
En México la raza Wagyu, fue introducida a través de embriones procedentes de Estados Unidos y actualmente se produce a través de cruzamientos con razas cárnicas con la intención de mejorar características de la canal y adaptabilidad a las condiciones de producción y actualmente, su carne es comercializada bajo el nombre Wagyu-Cross que utiliza la estrategia de cruzamientos con otras razas de ganado para carne para combinar las habilidades de conformación y tamaño de canal con la habilidad de marmoleo y deposición grasa 11, sin embargo, las ventajas relativas de esta estrategia no han sido evaluadas. Por lo tanto, el objetivo este estudio fue caracterizar y comparar la composición de FA en carne Wagyu y sus cruzas, y estudiar el efecto de un panel de marcadores genéticos en genes candidatos del metabolismo lipídico sobre la composición de los FA.
MATERIALES Y MÉTODOS
Origen de las muestras. Se colectaron 111 muestras de carne del músculo Longissimus dorsi de la sección entre la 11a y 12a costilla, de animales provenientes de la ganadería rancho "Exhacienda Cañas" ubicado en el municipio de Canatlán Durango, México, 24.806 N, -104.815 O, con clima semiárido, cálido con lluvias en verano.
Las muestras se agruparon de acuerdo al grupo genético en: Wagyu (WAG, n=39) y sus cruzas con Angus (WAN, n = 18), Beefmaster (WBM, n = 12), Brangus (WBR, n=29) y Hereford (WHF, n = 13). Los animales muestreados fueron alimentados durante el engorde con una dieta con formulación especial a base de maíz, heno, salvado, pasta de soya, granos secos, ensilado, alfalfa y calcio. Los animales tenían una edad promedio de 33.6±8.9 meses, y fueron engordados durante 428± 149.6 días en promedio y sacrificados en un rastros de Tipo Inspeccion Federal (TIF).
Cuantificación de ácidos grasos. Las muestras de carne se procesaron con el Lipid Extraction Kit No. Catálogo MAK174 (Sigma-Aldrich®, St. Louis, MO) utilizando el protocolo sugerido por el fabricante y solución de trifluoruro de boro-metanol para el proceso de transesterificación. La cuantificación de FA se realizó en un cromatógrafo de gases líquidos/espectometría de masas (GC/MS) GC7890A (Agilent Technologies, Santa Clara, CA), con la columna 123-2332 (Agilent Technologies, Santa Clara, CA) DB-23 (30 m x 0.32 mm, 0.25 um), siguiendo los parámetros previamente reportados 11, donde la temperatura de inyeccion fue de 50°C, seguido de incrementos de 10°C/min hasta alcanzar los 180°C donde se mantuvo por 5 min, posteriormente se utilizaron incrementos de 5°C/min hasta alcanzar los 220°C donde se mantuvo por 20 min. El tiempo de lectura fue de 48 min a una temperatura máxima de 240°C. El gas acarreador fue el helio con flujo total de 29.174 mL/min. La lectura de los cromatogramas se realizó con el programa Agilent G1701EA ChemStation GC/MSD (Agilent Technologies, Santa Clara, CA), tomando como base el tiempo de retención de los FA del standard Supelco® 37 Component FAME mix (Supelco®, Burlington, MA). Las concentraciones de FA en las muestras (g/100g de FA), se obtuvieron de los porcentajes de las áreas de los picos cuantificados 11.
Genotipificación. Se seleccionó un panel de 20 SNPs localizadods en 17 genes candidatos para determinar su asociación con la deposición de FA tomando como base el diseño de Vela 11. Los SNPs se seleccionaron considerando su participación en alguna ruta metabólica asociada a los FA y su efecto reportado en la composición de los FA en carne o leche (Tabla 1). Se realizó la extracción de ADN de las muestras con el kit comercial Genelute Mammalian Genomic DNA Cat. G1N350 (Sigma-Aldrich®, St. Louis, MO). La tipificación se realizó en la compañía NEOGEN Corp. con la tecnología Sequenom massARRAY® system (Agena Bioscience, San Diego, CA).
Tabla 1 Panel de 22 SNPs seleccionados para la asociación a ácidos grasos.
| Gen | Cromosoma | Locus | Alelos | Ref |
|---|---|---|---|---|
| ACACA | 19 | ss64381883 | G/A | 12 |
| DGAT1 | 14 | ss77831745 | G/A | 13 |
| HNF4A | 13 | ss61961144 | T/C | 12 |
| IGF2R | 9 | ss77831885 | G/A | 12 |
| INSIG2 | 2 | rs134478878 | C/G | 14 |
| INSIG2 | 2 | rs133396141 | C/T | 14 |
| LPL | 8 | ss65478732 | T/C | 12 |
| MEF2C | 7 | ss38329156 | G/T | 12 |
| MYOZ1 | 28 | ss77831945 | T/C | 12 |
| PLTP | 13 | ss77832104 | G/A | 12 |
| PPARG | 22 | ss62850198 | A/G | 12 |
| PPARGC1A | 6 | c.1847 | C/T | 15 |
| PPARGC1A | 6 | c.1892 + 19 | T/C | 15 |
| PRKAR2A | 22 | ss62837580 | T/C | 12 |
| PRKAR2A | 22 | ss62837667 | T/C | 12 |
| SCD | 26 | c.702 | A/G | 16 |
| SCD | 26 | c.762 | T/C | 16 |
| SCD | 26 | c.878 | C/T | 16 |
| SCD5 | 6 | rs109937152 | C/T | 14 |
| SLC2A4 | 19 | ss62538460 | G/A | 12 |
| SREBF1 | 19 | ss62543518 | T/C | 12 |
| SRPR | 29 | rs110036978 | C/G | 14 |
Análisis estadísticos. Se examinó el equilibrio de Hardy-Weinberg (HW) de cada locus estudiado por grupo genético, adicionalmente el desequilibrio de ligamiento utilizando el software GENEPOP 17.
Para estimar el efecto de las cruzas de Wagyu y ajustar la concentración de FA, para la posterior asociación con los genotipos en los loci estudiados, se ajustó un modelo mixto utilizando el procedimiento MIXED, de la siguiente manera:
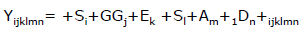
Donde: Yijklmn= variable aleatoria de concentración de FA, = media general, Si= efecto aleatorio del semental, GGj= efecto fijo del j-ésimo Grupo Genético, Ek= efecto fijo del k-ésima extracción de FA, Sl= efecto fijo del l-ésimo sexo del animal, Am= efecto fijo del m-ésimo año, Dn= Efecto fijo de la covariable lineal de los m-ésimo día en engorde, y ijklmn= error aleatorio residual, considerado posteriormente como la variable aleatoria de ajuste de la concentración de FA en carne (aYi). Las medias de mínimos cuadrados de cada Grupo Genético fueron estimadas y se realizó una comparación de medias con una prueba de t de mínima diferencia significativa y considerando un alfa= 0.05.
Posteriormente, se ajustó un modelo lineal poligénico para estimar el efecto de los genotipos en los loci estudiados de la siguiente manera:
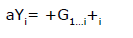
Donde: aYi= Variable aleatoria ajustada de concentración de FA, = media general, Gi= efecto poligénico fijo de los n= 1...¡ loci y i= error aleatorio residual. Las medias de mínimos cuadrados de los genotipos en cada locus fueron estimadas y se realizó una comparación de medias con una prueba de t de mínima diferencia significativa con ajuste de Bonferroni para múltiples comparaciones, considerando un alfa= 0.05.
Todos los procedimientos estadísticos fueron realizados utilizando el software SAS v.9.4 (SAS Institute Inc., Cary, CN).
RESULTADOS
Composición de los ácidos grasos. Se determinó la composición de los siguientes FA: láurico (C12:0) mirístico (C14:0), miristoleico (C14:1), pentadecanoico (C15:0), palmítico (C16:0), palmitoleico (C16:1), heptadecanoico (C17:0), heptadecenoico (C17:1), esteárico (C18:0), oleico (18:1), linoleico (C18:2), a-linolénico (C18:3), eicosanoico (C20:0), eicosenoico (C20:1), eicosadienoico (C20:2) y eicosatrienoico) (C20:3), además de las proporciones MUFA/SFA y PUFA/SFA de carne Wagyu y Wagyu-cross. Las medias de FA de la población se presentan en la tabla 2.
Tabla 2 Perfil de AG (g/100g de carne) en la población Wagyu y cruzas. n=83.
| Ácido graso | Símbolo | Media±EE |
|---|---|---|
| Ácido Laurico | C12:0 | 0.120±0.075 |
| Ácido Miristico | C14:0 | 3.759±1.076 |
| Ácido Miristoleico | C14:1 | 0.746±0.408 |
| Ácido Pentadecanoico | C15:0 | 0.466±0.210 |
| Ácido Palmitico | C16:0 | 27.509±5.713 |
| Ácido Palmitoleico | C16:1c | 3.710±1.505 |
| Ácido Heptadecanoico | C17:0 | 0.837±0.377 |
| Ácido Heptadecenoico | C17:1 | 0.467±0.253 |
| Ácido Esteárico | C18:0 | 16.731±6.122 |
| Ácido Oleico | C18:1 | 40.877±6.433 |
| Ácido Linoleico | C18:2 | 4.348±1.608 |
| Ácido a-Linolénico | C18:3 | 0.218±0.100 |
| Ácido Araquídico | C20:0 | 0.160±0.140 |
| Ácido Eicosenoico | C20:1 | 0.289±0.131 |
| Ácido Eicosadienoico | C20:2 | 0.102±0.138 |
| Ácido Dihomo-y-linolenico | C20:3 | 0.183±0.171 |
| Ácidos grasos saturados | ΣSFA | 49.421±5.905 |
| cidoss grasos monoinsaturados | ΣMUFA | 45.801±6.393 |
| Ácidos grasos poliinsaturados | ΣPUFA | 4.776±1.922 |
| MUFA/SFA | 0.952±0.224 | |
| PUFA/SFA | 0.097±0.040 |
Se analizaron las medias de mínimos cuadrados del efecto de las cruzas sobre la deposición de FA en el longissimus dorsi de los grupos WAN, WBM, WBR, WHF y WAG (Tabla 3). WBM presentó la mayor concentración de C16:0 y C18:0 y ΣSFA, donde se observó una diferencia (5.3%, 6.2% y 13.0%, respectivamente) respecto a WBR, el cual fue el grupo con la menor concentración, no obstante, no se obtuvieron diferencias significativas (p≥0.05) entre los grupos. Además, C15:0 y C17:0 se encontraron en mayor concentración en WBM, pero solo para C17:0 se encontró diferencia significativa respecto a los demás grupos genéticos (p<0.01) estudiados. Por otro lado, WBM tuvo la menor concentración del C18:1, ΣMUFA y C18:2, donde se observó una diferencia (10.213%, 11.039%, y 2.754%, respectivamente), respecto a WBR, sin embargo, no se obtuvieron diferencias significativas (p≥0.05).
Tabla 3 Medias de mínimos cuadrados del efecto de la cruza con ganado Wagyu sobre la deposición de AG en carne.
| FA | WAN | WBM | WBR | WHF | WAG | P-value |
|---|---|---|---|---|---|---|
| n= 18 | n= 12 | n= 23 | n = 12 | n= 31 | ||
| C12:0 | 0.217±0.046 | 0.161±0.071 | 0.249±0.045 | 0.183±0.046 | 0.190 ±0.040 | 0.1312 |
| C14:0 | 3.402±0.918 | 3.012±1.408 | 2.857±0.888 | 3.352±0.901 | 2.971±0.802 | 0.8433 |
| C14:1 | 0.492±0.311 | 0.602±0.477 | 0.329±0.301 | 0.654±0.305 | 0.510±0.271 | 0.7473 |
| C15:0 | 0.402±0.128 | 0.730±0.197 | 0.379±0.124 | 0.364±0.126 | 0.367±0.112 | 0.4437 |
| C16:0 | 28.380±3.506 | 31.604±5.376 | 26.279±3.391 | 28.056±3.442 | 28.363±3.061 | 0.7128 |
| C16:1 | 2.706±0.911 | 1.895±1.398 | 3.075±0.881 | 2.591±0.895 | 2.623±0.796 | 0.8519 |
| C17:0* | 0.810±0.225b | 1.890±0.345a | 0.819±0.218b | 0.534±0.221b | 0.689±0.196b | 0.0026 |
| C17:1 | 0.490±0.121 | 0.601±0.186 | 0.522±0.117 | 0.379±0.119 | 0.457±0.106 | 0.4559 |
| C18:0 | 18.621±4.185 | 24.081±6.418 | 17.928±4.048 | 19.157±4.109 | 18.519±3.655 | 0.9395 |
| C18:1 | 39.849±4.810 | 31.543±7.377 | 41.756±4.653 | 41.095±4.722 | 40.281±4.201 | 0.6626 |
| C18:2 | 4.376±1.286 | 2.848±1.973 | 5.602±1.244 | 3.502±1.263 | 4.741±1.123 | 0.3482 |
| ΣSFA | 51.834±4.239 | 61.481±6.501 | 48.514±4.100 | 51.648±4.162 | 51.101±4.162 | 0.3352 |
| ΣMUFA | 43.538±4.620 | 34.643±7.085 | 45.682±4.469 | 44.721±4.536 | 43.873±4.034 | 0.5524 |
| MUFA/SFA | 0.826±0.164 | 0.528±0.251 | 0.935±0.158 | 0.843±0.162 | 0.862±0.143 | 0.5406 |
| PUFA/MUFA | 0.084±0.0322 | 0.052±0.049 | 0.118±0.310 | 0.064±0.031 | 0.095±0.028 | 0.3101 |
FA: Ácido graso. *p<0.05. WAN: Wagyu x Angus. WBM: Wagyu x Beefmaster. WBR: Wagyu x Brangus. WHF: Wagyu x Hereford. WAG: Wagyu. SFA: Ácidos grasos saturados. MUFA Ácidos grasos monoinsaturados PUFA Ácidos grasos poliinsaturados
Análisis del efecto de los marcadores en genes candidatos sobre la concentración de los ácidos grasos de carne bovina. De los SNPs analizados, se encontró que HNF4A ss61961144 y SCD c.762 se encontraron en desviación del equilibrio de HW (p<0.001 y p<0.05, respectivamente). Por otra parte, se encontró que SCD c.702 se encuentra en desequilibrio de ligamiento con SCD c.762, asimismo SCD c.702 y SCD c.762 mostraron ligamiento a SCD c.878 (p<0.001); de igual manera que los SNPs ss62837667 y ss62837580 ubicados en el gen PRKAR2A mostraron estar en desequilibrio de ligamiento (p<0.001).
Se encontraron nueve marcadores con efecto sobre la composición de FA en carne Wagyu y Wagyu-Cross (Tabla 4). Dentro de los genes que interactúan en la ruta catabólica de la proteína quinasa activada por AMP (AMPK), el marcador MEF2C ss38329156 se encontró asociado a C14:0 (p<0.05), PPARGC1A c.1892+19 a C18:2 (p<0.05) y SLC2A4 ss62538460 a ΣSFA (p<0.01), ΣMUFA y MUFA/SFA (p<0.05). En el conjunto de genes analizados que participan en la ruta anabólica AMPK, los tres marcadores en SCD se asociaron a diferentes FA. El marcador c.702 se asoció a C14:0 y C16:0 (p<0.05) C762 a ΣSFA (p<0.05) y c.878 a ΣSFA (p<0.01), C16:0, C17:1, C18:1, ΣMUFA y MUFA/SFA (p<0.05). El marcador PLTP ss77832104 fue el único de los genes analizados en la ruta del colesterol que tuvo una asociación, este marcador tiene efecto sobre C16:0 (p<0.05). Finalmente, en el resto de los marcadores analizados IGF2R ss77831885 se asoció a C16:0 (p<0.05) y MYOZ1 ss77832104 a C17:1 (p<0.05).
Tabla 4 Efecto de polimorfismos en genes candidatos sobre la concentración de ácidos grasos de carne bovina
| Gen y locus | A/B | AAF | FA | AA | MMC ± EE AB | BB | P-value |
|---|---|---|---|---|---|---|---|
| MEF2C ss38329156 | G/T | 0.784 | C14:0 | -2.783±1.761a | -1.367±1.614b | -3.010±2.030ab | 0.0367 |
| PPARGC1A c.1892+19 | T/C | 0.194 | C18:2 | -2.003±3.258ab | -1.751±2.645a | 0.204±2.413b | 0.0353 |
| SLC2A4 ss62538460 | G/A | 0.969 | ΣSFA | -4.864±2.413b | -14.537±8.975a | - | 0.0090 |
| ΣMUFA | 5.276±7.686b | 14.922±8.975a | - | 0.0350 | |||
| AGM/AGS | 0.204±0.266b | 0.570±0.311a | - | 0.0224 | |||
| SCD c.702 | A/G | 0.662 | C14:0 | -1.844±1.606ab | -0.587±1.564b | -4.730±1.564a | 0.0140 |
| C16:0 | -0.004±5.072b | -0.263±4.941b | -12.002±7.366a | 0.0322 | |||
| SCD c.762 | T/C | 0.680 | ΣSFA | -6.477±6775ab | -6.913±6.590b | -15.711±6.778a | 0.0202 |
| SCD c.878 | C/T | 0.0.593 | C16:0 | -7.358±5.995a | -5.891±6.04a | 0.978±5.052b | 0.0270 |
| C17:1 | -0.104±0.229b | 0.2151±0.23a | 0.018±0.193ab | 0.0154 | |||
| C18:1 | 16.620±9.800ab | 15.001±9.876a | 3.450±8.259b | 0.0255 | |||
| ΣSFA | -14.598±6.950a | -11.859±7.003a | -2.643±5.857b | 0.0087 | |||
| ΣMUFA | 14.325±8.897ab | 13.212±8.966a | 2.759±7.498b | 0.0272 | |||
| AGM/AGS | 0.545±0.308ab | 0.484±0.311a | 0.133±0.260b | 0.0302 | |||
| PLTP ss77832104 | G/A | 0.785 | C16:0 | -8.638±5.856a | -6.060±5.679a | 2.427±5.8052b | 0.0394 |
| IGF2R ss77831885 | G/A | 0.829 | C16:0 | -5.477±5.834ab | -7.730±5.795a | 0.936±5.652b | 0.0423 |
| MYOZ1 ss77832104 | T/C | 0.396 | C17:1 | -0.123±0.202b | 0.061±0.218ab | 0.181±0.226a | 0.0427 |
A/B: Relación alélica de locus. AAF: Frecuencia de alelo A, FA: Ácido graso.
DISCUSIÓN
La carne Wagyu es reconocida mundialmente por desarrollar carne con alto contenido de IMF, esta es una de las características más importantes debido a que mejora las percepciones sensoriales como sabor, jugosidad y suavidad 18,19; además, el contenido y proporción de los FA en las carnes rojas se ha relacionado a efectos en la salud.
Generalmente los SFA son los más abundantes de la carne bovina y son asociados negativamente a la salud; C16:0 y C18:0 son los SFA más abundantes en la carne bovina, en la población analizada la suma de estos equivale al 89.5% de los cuales C12:0, C16:0 y C18:0 se han asociado a riesgo de diabetes tipo II, cardiopatía coronaria y enfermedades cardiovasculares 7,20. Por otro lado, C16:1 y C18:1 son los MUFA más abundantes y ambos se han asociado a mejorar la percepción sensorial en la carne bovina 8,21. En conjunto estos MUFA conforman el 97.3% de ΣMUFA. Los PUFA se han relacionado con múltiples efectos positivos en la salud 22,23. Debido a lo anterior se busca mejorar las proporciones MUFA/SFA y PUFA/SFA en la carne. No existe un consenso sobre la proporción ideal de MUFA/SFA, sin embargo, esta proporción permite comparar la composición de grasa. En la raza Wagyu, generalmente se reportan valores de 1.17 9,10, en este estudio, se encontró un valor de 0.95. La relación PUFA/SFA es uno de los principales parámetros que se utiliza para medir la calidad nutricional de la proporción lipídica de los alimentos. Las pautas nutricionales recomiendan una relación superior a 0.4, la proporción para la carne bovina es cercana a 0.1 24, en el presente estudio se encontró un valor de 0.097.
Diferencias en la deposición de los ácidos grasos de Wagyu y sus cruzas. La diferencia genética de las razas influye sobre el metabolismo de los FA provenientes de fuentes dietarias y la síntesis de novo, los cuales influyen en gran medida en el contenido de SFA y MUFA 22,25,26. En la tabla 3, se presentan las medias de mínimos cuadrados de la concentración de FA en el Longissimus de las diferentes cruzas estudiadas. De los grupos analizados WBR obtuvo la mejor proporción de FA, que aumentó tanto la relación MUFA/SFA como PUFA/SFA, sin embargo, sin diferencias significativas (p≥0.05). En los resultados obtenidos solo C17:0 tuvo diferencia significativa donde WBM produjo hasta 3.5 veces más contenido, en contraste con WHF. Estudios de población a gran escala han demostrado efectos de C17:0 sobre la reducción de riesgos de mortalidad, obesidad y numerosas enfermedades cardio-metabólicas y hepáticas 7,27.
Estudios previos han dejado claro la diferencia que existe entre las razas Angus y Wagyu. En novillos Wagyu y Angus alimentados con una dieta especial (ej. Japonesa), se encontraron diferencias significativas donde el músculo longissimus dorsi de Wagyu tuvo mayor concentración de C14:1 y menor de C18:0 con mayor proporción MUFA/SFA, además en la IMF y grasa subcutánea se encontraron diferencias en cuatro FA que influyeron favorablemente sobre la concentración MUFA/SFA de Wagyu 9. En novillos Wagyu y Angus se encontraron diferencias significativas en seis FA del longissimus; tres favorecían la proporción MUFA/SFA de Wagyu y dos la proporción de Angus, no obstante, no se observaron diferencias significativas en MUFA/ SFA. Angus presentó mayor concentración en C18:2, lo que mejoró significativamente la proporción PUFA/SFA. Además, se encontraron diferencias de seis FA de grasa subcutánea 28. En otro análisis donde se evaluaron distintos sistemas de finalización entre ganado Angus y Wagyu se encontraron diferencias por efecto de la raza en C14:0, C16:0 y tres PUFA de grasa subcutánea, donde los animales Wagyu presentaron menor concentración de los C14:0 y C16:0 y mayor concentración de PUFA en la mayoría de los sistemas de finalización 29.
Existen pocas comparaciones de FA en carne de cruzas Wagyu, respecto a otras razas. En animales Angus y sus cruzas con Brangus y Brahman (25%, 50% y 75%, respectivamente), se reportaron diferencias de múltiples FA en el longissimus. En los grupos Angus 25%, Angus 50% y Brahman 75% obtuvieron las menores concentraciones de SFA debido a que también tenían menor concentración de C16:0, adicionalmente el grupo Brahman mostró mayor concentración de 9 PUFA reflejando una diferencia en ΣPUFA 20. En cruzas Hereford con Angus y Limousine se encontró diferencia de C16:0 ΣSFA, ΣMUFA, 7 PUFA 30. Otro estudio de cruzas Hereford, con Belgian Blue, Limousine y Rubia gallega, reveló diferencias entre estas razas de los FA C10:0 y C18:0 31.
Análisis del efecto de los marcadores en genes candidatos sobre la concentración de los ácidos grasos de carne bovina. La promesa de mejoramiento genético mediante marcadores moleculares ha incentivado el descubrimiento y validación de los marcadores genéticos que incluye estudios en diferentes razas bovinas. En el presente estudio, se encontraron nueve marcadores genéticos con efecto sobre la composición de FA en carne Wagyu y Wagyu-Cross (Tabla 4).
Algunos de los marcadores que mostraron efectos significativos forman parte de la vía AMPK, que participa como sensor del estado energético celular activado por aumentos de la proporción celular AMP/ATP causado por estrés metabólico que interfiere con la producción de ATP. Una vez activada, la vía AMPK induce rutas catabólicas como la oxidación y la glucólisis de FA para la producción de ATP 32. En la vía de señalización AMPK, PPARG es regulado por PPARGC1A y este induce a SLC2A4, el cual activa la biogénesis mitocondrial; por otra parte, MEF2 interacciona específicamente al elemento de respuesta MEF2C del gen SLC2A4, que es esencial para la expresión de GLUT4 en el músculo esquelético 12.
El gen PPARGC1A sintetiza el Coactivador del receptor gamma 1-alfa activado por el proliferador de peroxisomas, en el intrón 9 de este gen, se encuentra la transición c.1892+19, donde el alelo T de este marcador, se asoció al incremento de grasa en leche de ganado Holstein 15,33. En este análisis se encontró que el alelo T redujo la concentración de C18:2. El homocigoto CC produjo 1.955 g/100g más que TC (p<0.05). PPARGC1A regula la expresión de genes implicados en la adipogénesis, gluconeogénesis, el metabolismo oxidativo y termogénesis adaptativa, homeostasis energética, además activa múltiples receptores de hormonas nucleares 15. Los ácidos linoleicos conjugados (CLA) conforman una familia de isómeros de 18 carbonos y dos dobles enlaces (C18:2) los cuales han sido asociados a efectos anticancerígenos, anti-obesidad y antidiabéticos 34. Por otra parte, el ácido linoleico (C18:2c9,12) es un co-6 esencial debido a que no puede ser sintetizado por humanos, este es el PUFA más abundante en la carne y ha sido correlacionado positivamente a sabor, jugosidad y suavidad 21.
El gen MEF2C sintetiza al Factor potenciador de miocitos 2C, en el intrón 10 se encuentra la transversión ss38329156, donde el homocigoto GG se asoció 50% con menor concentración de C14:0 respecto al heterocigoto GT (p<0.05). Este dato respalda lo encontrado previamente 11 donde GG se asoció a menor concentración de C14:0, C15:0, C16:0, C16:0 C17:0 en carne comercial mexicana, incluida Wagyu-Cross. PPARGC1A y MEF2C son genes activos del músculo esquelético involucrados en el metabolismo de las grasas y el desarrollo muscular. Sin embargo, MEF2C se expresa selectivamente en miocitos diferenciados y activa casi todos los genes del músculo esquelético y cardíaco, controlando la morfogénesis, la miogénesis cardíaca y el desarrollo vascular 12.
En el exón 11 de SLC2A4, se encuentra la transición ss62538460, en los presentes resultados, el genotipo GA de este SNP se asoció a un 66% menor contenido de SFA y 64% mayor de MUFA y MUFA/SFA (p<0.001, p<0.05 y p<0.05, respectivamente) respecto a GG. Previamente este marcador se asoció a C17:0 y FA totales 11. La proporción de FA influye sobre los FA totales, por lo que la previa asociación puede estar validada por los resultados aquí obtenidos. Es posible que el alelo A tenga un efecto aditivo, sin embargo, no se encontraron homocigotos. Al igual que en esta población, en razas españolas se encontró baja segregación del alelo A 15. El Miembro 4 de la familia 2 de transportadores de solutos (GLUT4) es una proteína producida por SLC2A4 la cual capta la glucosa mediada por insulina en células musculares y adiposas 12. Las diferencias en el impacto fisiológico de la expresión de SLC2A4 en el desarrollo adiposo de los rumiantes podría deberse al hecho de que la glucosa es un sustrato menor para la síntesis de FA 12.
Además de activar rutas catabólicas, la vía AMPK inhibe procesos anabólicos como la síntesis de proteínas, FA y glucógeno para restablecer el equilibrio energético 12. La vía AMPK inhibe al SREBF1, el cual regula los genes ACACA y SCD, los cuales participan en vías anabólicas como la síntesis de los FA y la adición de dobles enlaces a estos 16. El gen SCD sintetiza la enzima Estearoil Co-A desaturasa, la cual cataliza la inserción de un doble enlace cis en la posición A-9 en sustrato. Taniguchi et al 16 descubrió tres SNPs del exón 5, los cuales se asociaron a distintos FA de este estudio (Tabla 4). Los datos muestran que el alelo A en c.702 aumenta la concentración de C14:0, el homocigoto GG se asoció a 87% menor concentración respecto al heterocigoto AG (p<0.001) además, el homocigoto GG se asoció a 11.998 g/100g menor concentración respecto a AA y 11.739 g/100 g respecto a AG de C16:0 (p<0.05). En el marcador c.762 la presencia del alelo T aumenta SFA, el homocigoto CC se asoció a 55% menor concentración, respecto al heterocigoto TC (p<0.05). En el marcador c.878, se observó un efecto aditivo del alelo C, los genotipos CC y CT se asociaron a menor concentración de C16:0 en 8.336 y 6.869 g/100g respecto a TT (p<0.05) y 11.955 y 10.453 en SFA (p<0.05); además, CT se asoció a 11.551g/100g, 10.453 g/100g y 0.351, menor concentración de C18:1, MUFA y MUFA/ SFA, respecto a (p<0.05); por otro lado, CT se asoció a 0.3191 g/100g mayor concentración de C17:1 respecto a CC (p<0.05). Múltiples estudios coinciden en la asociación del alelo C del marcador c.878 en el aumento de C18:1 y MUFA de distintas razas 15,16,35,36, sin embargo, no se han reportado efectos de C16:0 y C17:1. El efecto de este marcador sobre C16:0 se relaciona la enzima Estearoil Co-A desaturasa que utiliza a este FA como sustrato dando lugar a C16:1 12.
El colesterol forma parte de las membranas en las células de los mamíferos, así como un precursor de los ácidos biliares, la vitamina D y las hormonas esteroides. Sobre los genes que interactúan sobre el metabolismo del colesterol, la proteína de transferencia de fosfolípidos sintetizada por el gen PLTP transporta una gran cantidad de moléculas anfipáticas diferentes (como fosfolípidos, diacilglicerol y lipopolisacáridos), desempeñando una función fundamental en el metabolismo de lípidos y lipoproteínas 12. La transición ss77832104 ubicado en la región 3'UTR, ha sido asociado a la proporción n6/n3 en el músculo de razas españolas 37,38. En este estudio se observó un efecto aditivo en el alelo G; los genotipos GG y GA se asociaron a menor contenido (11.066 g/100g y 8.487 g/100g, respectivamente) de C16:0 respecto a AA (p<0.05).
Dentro del grupo sin relación con metabolismo de FA, IGF2R sintetiza el receptor 2 para el factor de crecimiento para la insulina el cual tiene múltiples funciones celulares como la diferenciación, migración y apoptosis de múltiples células 37. En estudios previos, el marcador ss77831885 se ha asociado a cinco FA y la intensidad de sabor de la carne de ganado español 37,38. En este estudio el heterocigoto GA presentó 8.666 g/100g menor concentración de C16:0 respecto a AA (p<0.05). Finalmente, el producto del gen MYOZ1 desempeña un papel importante como proteína de unión intracelular y localiza señales de calcineurina en el sarcómero 12. En razas españolas se encontró que el marcador ss77832104 influye sobre el CLA, la proporción C18:2/C18:3 y C22:6 37. En el presente análisis se observó un efecto aditivo del alelo C, el homocigoto CC presentó 0.303 g/100g mayor contenido de C17:1 respecto a TT (p<0.05).
En el presente estudio se evaluó la deposición de carne Wagyu y Wagyu-cross conocida por producir carne con alto contenido de IMF. Esta característica y la composición de FA influye sobre los conceptos de calidad sensorial y nutricional. De manera general se identificaron FA poliinsaturados que permiten confirmar la deposición favorable en la población estudiada, sin embargo, la distribución de las muestras no permitió en algunos casos su comparación. Los resultados mostraron que la carne Wagyu y Wagyu-Cross, tienen una composición de FA muy similar entre grupos evaluados, pero resaltó la diferencia 3.5 veces mayor concentración de C17:0 de WBM respecto a los demás grupos. Esto permite concluir que no hay un efecto importante en la deposición de los FA entre Wagyu y sus cruzas, pero que aspectos específicos en la deposición como en C17:0 podrían ser sujetos a estudios particulares para determinar su confirmación.
Concluyentemente, se encontraron 17 asociaciones significativas en 9 SNPs vinculados a la composición de FA en carne Wagyu y Wagyu-Cross, algunas asociaciones confirman reportes previos, sin embargo, la mayoría de las asociaciones encontradas en este estudio, son nuevas. Entre las nuevas asociaciones es importante destacar el efecto del marcador SLC2A4 ss62538460 donde el heterocigoto GA se asoció a la reducción considerable de las concentraciones MUFA, SFA y la proporción MUFA/SFA. Además, los marcadores, PLTP ss77832104 y IGF2R ss77831885 se asociaron a C16:0, el SNP MYOZ1 ss77832104 se asoció a C17:1 y PPARGC1A c.1892+19 a C18:2. Por otro lado, las asociaciones en MEF2C ss38329156 y SCD c.878 respaldan asociaciones descritas anteriormente. Estas evidencias confirman a SLC2A4, PLTP, MYOZ1, PPARGC1A y SCD como genes candidatos y a sus marcadores para su potencial utilización en estrategias de selección asistida. De esta manera esta información también permite ampliar el número de marcadores para el entendimiento de la arquitectura genética de la composición de FA de la carne bovina.
Finalmente, es importante resaltar que un mayor tamaño de muestra, que también implica aumento de costos de financiamiento, podrían validar estos resultados sobre todo considerando la evaluación de efectos correlacionados con los beneficios de otros caracteres de la canal (p.e. grasa intramuscular) con la deposición de FA en las cruzas derivadas de Wagyu y asegurar la mejor implementación en el uso de la información con fines comerciales y de los marcadores estudiados de manera integrada en programas de mejoramiento asistido.

