











 Spanish (pdf)
Spanish (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mail Cited by SciELO
Cited by SciELO  Cited by Google
Cited by Google  Similars in
SciELO
Similars in
SciELO  Similars in Google
Similars in Google 
 Permalink
Permalink
Introducción
A nivel mundial, existen dos especies de palma aceitera de gran importancia económica para la agroindustria: Elaeis guineensis Jacq. y Elaeis oleifera (HBK) Cortés. La primera especie es originaria del centro y occidente de África, mientras que la segunda, del norte de Sudamérica y Centroamérica; sin embargo, la mayoría de las plantaciones comerciales en el mundo están sembradas con material africano E. guineensis (Rey et al., 2004). Malasia e Indonesia son los mayores productores de palma aceitera y aportan entre el 80% y el 86% de la producción mundial del aceite crudo de palma (Mesa, 2018).
La industria de palma aceitera se desarrolló gracias a la utilización del híbrido Ténera de palma africana como principal material de siembra, proveniente del cruzamiento de los tipos Dura y Pisifera (Rajanaidu, 2017). En 1936 se inició una distribución de material genético desde el jardín botánico de Lacentilla, Honduras, para los países de América Tropical (Richardson, 1995). Los programas de mejoramiento Malaysian Palm Board (MPOB), el Instituto Francés IRHO/CIRAD y la empresa ASD Costa Rica, así como Indonesia, Colombia y Ecuador han realizado significativas colectas de germoplasma de palma africana y palma americana para la selección de nuevas variedades (Rajanaidu, 2017).
La semilla de palma aceitera es la nuez que queda después de la extracción del mesocarpio aceitoso y suave del fruto, y está conformada por cuesco o endocarpio y por una, dos o tres almendras (Hartley, 1983). Ellis et al. (1991) clasificaron las semillas de E. guineensis como categoría intermedia entre recalcitrante y ortodoxa, después de observar que, al almacenar las semillas de esta especie a temperaturas entre 0 °C y -20 °C, disminuyó su viabilidad, pero con un bajo contenido de humedad conservaron su viabilidad.
Después de madurar en el interior del fruto, las semillas generalmente entran en latencia, la misma que se rompe cuando se exponen las semillas a periodos prolongados de calor, humedad, oxígeno, escarificación, entre otros (Taiz & Zeiger, 2007). En África Occidental, la semilla de palma aceitera en condiciones naturales germina esporádicamente en varios años. Sin embargo, bajo un tratamiento térmico continuo con temperaturas entre 38 °C y 40 °C, la germinación tiene lugar en pocos meses (Hussey, 1958). Existen dos técnicas utilizadas comercialmente para la germinación de semillas de palma aceitera (E. guineensis): el método de calor seco y el de calor húmedo, siendo el primero el más usado a nivel comercial (Corley & Tinker, 2016). El calentamiento es el tratamiento común para debilitar el opérculo y permitir el inicio de la germinación (Kelanaputra et al., 2018). Con el método de calor seco, las semillas se exponen a dos imbibiciones y a un calentamiento a temperaturas entre 37 °C y 39 °C por 50 días (Mok, 1966, citado por Corley & Tinker, 2016). El método de calor seco tiene como ventaja que el contenido de humedad no es tan crítico como en el método de calor húmedo, ya que con una humedad relativa del 14-21% se puede lograr una germinación superior al 85% (Rees, 1962, citado por Corley & Tinker, 2016).
La función del ácido salicílico (AS) en la germinación de las semillas es controversial, ya que a veces induce la germinación y otras veces la inhibe, dependiendo de la dosis, la especie y el cultivar utilizado (Rodríguez-Larramendi et al., 2017). En maíz el AS estimula la germinación con dosis de 0,5-1,5 mM y la impide con dosis de 3-5 mM (Guan & Scandalios, 1995). Por otro lado, Fontana et al. (2017) informaron que, en semillas de Cichorium endivia L. (Asteraceae), el AS en concentraciones de 0,15-0,2 mM inhibe totalmente la germinación, mientras que con dosis menores el porcentaje de germinación es inferior al testigo. En semillas de Dracocephalum moldavica L. (Lamiaceae), una concentración de AS de 2 mM aumentó la germinación en condiciones de salinidad (Shaikh-Abol-Hasani, & Roshandel, 2019). Deenamo et al. (2018) observaron una reducción del 41% de afección por Phytophthora palmivora al realizar una aplicación exógena de AS en el follaje de plantas de caucho Hevea brasiliensis (Willd. ex A. Juss.) Müll. Arg. (Euphorbiaceae). Dzib et al. (2021) concluyeron que la imbibición por 24 h de las semillas de tomate (Solanum lycopersicum L.) (Solanaceae) en AS, utilizando dosis de 1 mM, 0,1 mM y 0,001 mM, no inhibe la germinación, favoreciendo las dosis más elevadas al crecimiento radicular en las plantas. Según Castillo.Méndez (2019), el AS estimula la germinación en Annona Muricata L. y ayuda a obtener plántulas más vigorosas.
El efecto protector en las plantas al estrés biótico y abiótico del aminoácido no proteico, ácido β-aminobutírico (BABA), se debe a la potenciación de mecanismos de defensa natural (Jakab et al., 2001). Al tratar las semillas de arroz Oryza sativa L. (Poaceae) con BABA, se observó en las plantas una reducción del estrés a la sequía y salinidad, lo que mejoró la actividad mitocondrial y fotosintética (Jisha & Puthur, 2016b). En otro estudio, estos mismos autores tuvieron resultados similares cuando trataron las semillas de Vigna radiata (L.) R. Wilczek (Fabaceae) con BABA (Jisha & Puthur, 2016a). Por otro lado, en Solanum betaceum Cav. (Solanaceae), el BABA redujo la lesión ocasionada por Phytophthora infestans (Castaño et al., 2015). Según Gur et al. (2021), el BABA induce resistencia contra Alternaria alternataf. sp. mali en frutos de manzana (Malus domestica Borkh).
La revisión de literatura científica demuestra la necesidad de evaluar la influencia de inductores de resistencia en la germinación de semillas de palma africana, con el propósito de encontrar alternativas que lleven a reducir el tiempo de germinación de semillas de palma aceitera africana y contrarrestar el daño ocasionado por patógenos durante el proceso de germinación.
Materiales y métodos
El estudio se llevó a cabo en la Estación Experimental Santo Domingo (EESD) del Instituto Nacional de Investigaciones Agropecuarias (INIAP), en la provincia Santo Domingo de los Tsáchilas, Ecuador, ubicada en las coordenadas X: -0,26608 y Y: -79,3802. Se utilizaron cuartos climatizados con temperatura y humedad relativa controlada para los tratamientos térmicos y la germinación. Los cuartos de calentamiento se programaron con una temperatura mínima de 38 °C y máxima de 40 °C, con una humedad relativa entre 35% y 45%. Los cuartos de germinación se programaron con temperatura entre 26 °C y 28 °C, y humedad relativa entre 45% a 65%. Se utilizaron semillas de tipo Dura, porque el grosor del cuesco de las semillas resultantes del cruzamiento Dura ´ Pisifera (Ténera) es similar al del parental femenino. El despulpado del fruto se realizó mecánicamente con un bingo despulpador; luego, las semillas fueron colocadas en zarandas bajo sombra a temperatura ambiente hasta alcanzar 15% de humedad. Se seleccionaron 30 semillas al azar por repetición y por cada tratamiento. Las semillas utilizadas tuvieron cero días de almacenamiento y no se utilizó fungicida para su preservación. Se utilizó un diseño completamente al azar (DCA) con arreglo factorial + 1, combinando cuatro factores: i) imbibición 1 (0, 2 y 6 días en agua + 2 días en inductor), ii) calentamiento (20 y 40 días), iii) imbibición 2 (0 y 2 días en agua + 2 días en inductor), y iv) inductores como AS y BABA en dosis de 1 mM y 2,5 mM, respectivamente. Esta combinación dio un total de 24 tratamientos más el testigo, que permaneció ocho y cuatro días en agua destilada durante la primera y segunda imbibición, respectivamente, y 80 días en calentamiento (tabla 1). Para la comparación de medias se utilizó la prueba de Tukey al 5%.
Tabla 1 Descripción de tratamientos mediante diferentes periodos de imbibición calentamiento e inductores sobre la germinación de la semilla de palma aceitera E guineensis Jacq

Fuente: Elaboración propia
Durante la primera imbibición, las semillas permanecieron cero, dos y seis días en agua destilada, para luego entrar dos días en solución de 1 mM de AS o 2,5 mM de BABA, de acuerdo con lo estipulado en la descripción de los tratamientos (tabla 1). Las semillas fueron secadas por 24 h a temperatura ambiente y, posteriormente, entraron a cuarto caliente por 20 y 40 días, respectivamente. Una vez extraídas las semillas del cuarto caliente, fueron llevadas a una segunda imbibición por cero y dos días en agua destilada, y dos días más en inductor, en dosis de 1 mM de AS y 2,5 mM de BABA. Las semillas entraron a un segundo secamiento por seis horas, para luego ingresar a cuartos de germinación. Con respecto al testigo, se utilizó la metodología aplicada por el INIAP para la producción de semillas de palma E. guineensis Jacq.
Después de 15 días de que las semillas entraron en cuarto de germinación, se evaluó cada semana el número de semillas germinadas, observando en la semilla el llamado punto blanco o inicio de germinación. Transcurrido 30 días de permanencia de las semillas en el cuarto de germinación, durante cuatro semanas consecutivas se identificaron las semillas germinadas con estándar óptimo, es decir, aquellas semillas cuyo embrión se desarrolló lo suficiente para identificar con facilidad la plúmula (longitud igual o superior a los 5 mm) y la radícula (longitud igual o superior a 10 mm), medición que se llevó a cabo con un calibrador y, al final del experimento, se contabilizaron las semillas germinadas de manera atípica o deforme en cada uno de los tratamientos.
En esta investigación se realizó un análisis estadístico de tipo descriptivo, haciendo uso del software Infostat para realizar el análisis de varianza (ANOVA, por sus siglas en inglés), análisis de media de todos los niveles de los factores y de sus interacciones, de interés para los investigadores, convirtiéndolo en un modelo con efecto fijo y con igual número de observaciones. Para la comparación de medias, se utilizó la prueba de Tukey al 5%. Además, se calcularon valores mínimos, máximos y desviación estándar en cada una de las variables, lo que sirvió para determinar el error estándar de las medias.
Resultados y discusión
Germinación de semillas
En el análisis estadístico para porcentaje de germinación de semillas, se encontró diferencia significativa entre los tratamientos y para el factor imbibición 1 (tabla 2).
Tabla 2 Valores significativos del ANOVA para germinación de semillas

**Altamente significativo Fuente Elaboración propia
Fuente: Elaboración propia
Al realizar las comparaciones de medias para imbibición 1, resultó que las semillas sometidas a ocho y seis días a remojo en esta fase fueron estadísticamente iguales entre sí y diferentes de aquellas que solo estuvieron dos días en remojo; la primera alcanzó 26,81% de semillas germinadas y la última 3,89% (figura 1). Azcón-Bieto y Talón (2008) informaron que la germinación es el proceso que se inicia con la toma de agua por la semilla seca. Por otra parte, Bewley y Black (2012) informaron que los procesos metabólicos de germinación temprana inician durante el remojo y que con el secado este proceso solo se detiene, pero no se revierte. Corrado y Wuidart (1990) recomendaron un remojo o imbibición por cinco días antes de que la semilla de E. guineensis Jacq. entre a calentamiento en el proceso de germinación.

Fuente: Elaboración propia
Figura 1 Comparación de medias y error estándar de porcentaje de germinación de imbibición 1 para el total de semillas germinadas
Cuando se realizó la comparación de medias, resultó que los tratamientos 13 y 9 fueron los que tuvieron mayor porcentaje de germinación con 41% y 39%, respectivamente, resultando estadísticamente iguales al testigo con una germinación de 26%. Nandeeshkumar et al. (2009) reportaron resultados similares en semillas de girasol, en donde los tratamientos con concentraciones de BABA de 10-50 mM) aumentaron la germinación y el vigor de las plántulas. Ambos tratamientos —13 y 9— tuvieron el mismo periodo de calentamiento y más de cuatro días de imbibición 1 (figura 2).

Fuente: Elaboración propia
Figura 2. Comparación de medias de porcentaje de germinación y error estándar de los tratamientos para el total de semillas germinadas. Medias con una letra en común no son significativamente diferentes (p > 0,05).
El porcentaje de germinación de los tratamientos y el testigo es inferior al promedio obtenido por grupo (20 racimos) en el INIAP. Sin embargo, dentro de los grupos procesados se han observado racimos con germinaciones óptimas inferiores a 50% y superiores a 90% (INIAP, 2018), lo que concuerda con Meerow y Broschat (1991), quienes informaron que la viabilidad de las semillas de palma puede variar entre plantas de la misma especie, incluso entre años en el mismo árbol. Las semillas utilizadas en esta investigación provienen de un racimo de polinización libre y de una planta con alta carga productiva, lo que coincide con Herrera y Alvarado (2012), quienes postulan que las semillas provenientes de plantas con alta carga productiva presentan menor porcentaje de germinación que aquellas que tienen menor carga productiva.
En la tabla 3 se indican los valores medios, mínimos, máximos y la desviación estándar (DE) de los tratamientos que presentaron diferencias estadísticas, además del testigo. Los tratamientos 9 y 13 presentaron un máximo de germinación de 47% y un mínimo de 23% y 33%, respectivamente; por otra parte, los tratamientos 5, 6 y 18 no superaron el 4% de germinación mínima y máxima, mientras que el testigo presentó un porcentaje de germinación mínimo de 20% y máximo de 33%.
Tabla 3 Valores medios mínimos máximos y desviación estándar de los tratamientos que presentaron diferencias estadísticas para germinación de semillas
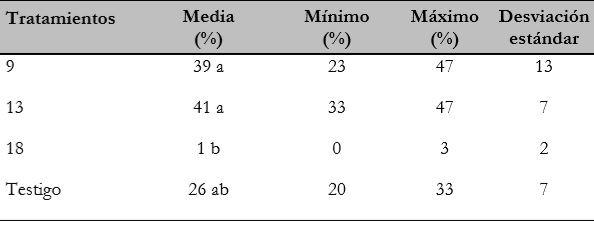
Fuente: Elaboración propia
Todos los tratamientos que permanecieron seis días en agua, más dos días en inductor durante la primera imbibición, con 20 días de calentamiento utilizando AS o BABA, iniciaron su germinación 47 días después de iniciado el proceso de germinación. Asimismo, a los 54 días de iniciado el proceso de germinación, todos los tratamientos que se sometieron en la primera imbibición a cuatro y seis días en agua más dos días de inductor mostraron germinación. Los tratamientos expresaron su mayor porcentaje de germinación a los 61 días de iniciado el proceso de germinación. La culminación de la germinación en casi todos los tratamientos que estuvieron 20 días de calentamiento se dio a los 75 días después de iniciado el proceso; es decir, la germinación se dio en un tiempo máximo de cuatro semanas (figura 3), a diferencia de lo reportado por Herrera et al. (1998), en donde la germinación se extendió por seis semanas con el uso de cianamida hidrogenada.

Fuente: Elaboración propia
Figura 3 Germinación de semillas de palma africana en el tiempo (47, 61, 75 y 82 días después de iniciado el proceso de germinación), de acuerdo con la combinación del total de días en imbibición 1 (8, 4 y 2 días) más el total de días en imbibición 2 (4 y 2 días) con inductor (AS o BABA), con un periodo de calentamiento de 20 días
Semillas germinadas dentro del estándar óptimo
Esta variable se definió seleccionando las semillas germinadas diferenciadas con plúmula y radícula bien desarrollada, con una longitud mínima de 0,5 cm de plúmula y de 1 cm de radícula; Kelanaputra et al. (2018) informaron que las longitudes máximas de radícula y plúmula aceptables para la venta son 2,0 cm y 1,5 cm, respectivamente. El análisis estadístico presentó valores altamente significativos para los tratamientos y valores significativos para la interacción imbibición 1 × calentamiento y para los factores vs. testigo (tabla 4).
Tabla 4 Valores significativos del ANOVA para porcentaje de semillas germinadas óptimas

*Significativo
**Altamente significativo
Fuente: Elaboración propia
Al analizar los inductores por separado en la interacción calentamiento × imbibición 1, se observó que para los tratamientos con AS y BABA el porcentaje medio de germinación óptima en esta misma interacción fue de 23,89% y 34,44%, respectivamente (figura 4), coincidiendo con lo expresado por Bermúdez-Ruiz et al. (2017) quienes, al someter las semillas de Bactris guineensis (L.) H.E. Moore (Arecaceae) a cinco días de imbibición, determinaron que estas alcanzaron la humedad suficiente para promover la germinación. Zambrano (1991) logró germinaciones de semillas de palma E. guineensis Jacq. superiores al 60%, con ocho días de imbibición antes de ingresar a calentamiento, y determinó que los periodos de calentamiento no deberían ser menores a 40 días, ni superiores a 80 días para obtener mayor porcentaje de germinación. Green et al. (2013) obtuvieron resultados diferentes a los presentados en esta investigación, concluyendo que el porcentaje de germinación máximo, dependiendo de los cultivares, se obtienen con periodos de calentamiento entre 45 y 80 días. Asimismo, Fondom et al. (2010) y Martine et al. (2009) coinciden en que 60 días de calentamiento de las semillas de palma es óptimo para la germinación.
Al realizar una prueba de comparación de medias para los tratamientos, al igual que en la variable de porcentaje de semillas germinadas, resultó que el tratamiento 13 es el que presenta mayor porcentaje de semillas germinadas diferenciadas dentro del estándar óptimo (40%), siendo este resultado menor en comparación con el 85% obtenido por Rees (1962, citado por Corley y Tinker, 2016) y Kelanaputra et al. (2018) en la producción comercial de semillas de palma, mediante el método de calor seco y con el uso de fungicidas para el control de patógenos; sin embargo, el tratamiento 13 es estadísticamente diferente a los tratamientos 5, 6, 11, 12, 17, 18, 20, 23 y al testigo (figura 5). Estos resultados coinciden con Nandeeshkumar et al. (2009), quienes observaron resultados similares en semillas de girasol, en donde los tratamientos con concentraciones de 10-50 mM de BABA aumentaron la germinación y el vigor de las plántulas. Por otra parte, Rajjou et al. (2006), en semillas de Arabidopsis thaliana (L.) Heynh. (Brassicaceae), observaron que el AS en concentraciones inferiores a 1 mM mejora la germinación en esta especie. Soare et al. (2018) reportaron que la tasa de germinación del cultivar AS de trigo mejoró al pretratar las semillas con concentraciones de 0,75 mM del inductor.

Fuente: Elaboración propia
Figura 4 Efecto del AS y BABA para la combinación de imbibición 1 (0, 2 y 6 días en agua más dos días en AS o BABA) × 20 y 40 días de calentamiento en la germinación de semillas germinadas óptimas, mediante líneas de tendencias calculadas en Excel.
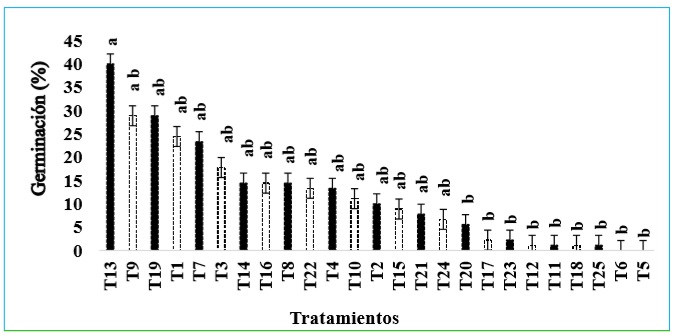
Fuente: Elaboración propia
Figura 5 Comparación de medias y error estándar de los tratamientos para germinación de semillas óptimas. Medias con una letra en común no son significativamente diferentes (p > 0,05).
En la tabla 5 se observa que el tratamiento 13 presentó valores mínimos y máximos de germinación con estándar óptimo, superiores al resto de los tratamientos con los que tuvo diferencias estadísticas, incluido el testigo, y con una media de 40% (± 5,77).
Tabla 5 Valores de media mínimo máximo y desviación estándar para los tratamientos que presentaron diferencias estadísticas para germinación de semillas con estándar óptimo
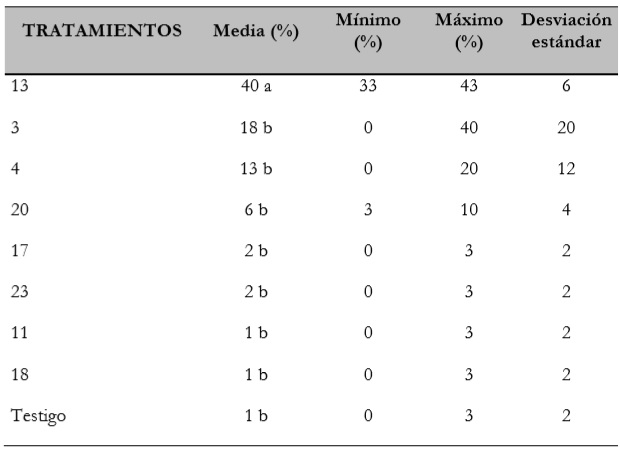
Fuente: Elaboración propia
La comparación de medias de los factores vs. testigo determinó que, en la variable de porcentaje de semillas germinadas óptimas, los factores superaron al testigo (figura 6). El testigo resultó tener mayor porcentaje de incidencia de efermedades (79%) con respecto a los factores que tuvieron una media de incidencia de 16% (figura 7), presentando en su mayoría contaminación con hongos. Carrillo et al. (2015) informaron que la germinación de las semillas de palma de aceite se ve afectada por una enfermedad denominada germen pardo, que puede ser causada por hongos de los géneros Aspergillus, Fusarium . Penicillium, que a su vez pueden actuar solos o en asociación entre ellos. Los resultados de la investigación concuerdan con lo informado por Jakab et al. (2001), quienes indican que BABA es un aminoácido no proteico, considerado un inductor abiótico, capaz de inducir resistencia a diversos patógenos en diferentes especies vegetales, y con Gurgel et al. (2005), quienes encontraron que, con aplicaciones foliares de BABA en tomate, se controlan problemas de Fusarium. Entretanto, el BABA aumentó la actividad enzimática en plantas de arándanos (Vaccinium corymbosum L.), logrando que esta especie presente mayor resistencia a la mancha foliar (Pestalotiopsismicrospora) (Yi-Lan et al., 2021). Durner et al. (1997) informaron que el ácido salicílico está involucrado tanto en las reacciones de defensa local en los sitios de infección, como en la reducción de resistencia sistémica, y que su síntesis y acumulación son importantes en la respuesta de defensa de las plantas. Farahani y Taghavi (2017) encontraron que el BABA en pimienta induce resistencia a Xanthomonas euvesicatoria.

Fuente: Elaboración propia
Figura 6 Comparación de medias y error estándar de porcentaje de germinación de factores vs. testigo para semillas germinadas óptimas. Medias con una letra en común no son significativamente diferentes (p > 0,05).

Semillas germinadas atípicas o deformes
Al momento de realizar la selección de semillas germinadas para la comercialización, se descartan aquellas semillas que están fuera de tipo, siendo las más comunes semillas atrofiadas, con dos raíces y con raíz larga (Kelanaputra et al., 2018). El análisis estadístico no mostró diferencias estadísticas para los tratamientos ni para factores vs. testigo referente a esta variable. Esto coincide con Shailasree et al. (2001), quienes utilizaron concentraciones de 25 mM y 50 mM de BABA en semillas de Pennisetum glaucum (L.) R. Br. (Poaceae) y reportaron que no hubo problemas en la germinación; en cambio, cuando usaron 75 mM y 100 mM de BABA, observaron inhibición en la germinación y en el vigor de las plántulas. Nandeeshkumar et al. (2009) observaron que concentraciones de BABA menores a 50 mM aumentó la germinación y el vigor de las plántulas. Entretanto, a concentraciones más altas hubo un efecto contrario. Finalmente, Rajjou et al. (2006) observaron que en semillas de Arabidopsis thaliana la germinación y el desarrollo de las plántulas se vieron afectados negativamente.
Conclusiones
Las semillas de palma aceitera (E. guineensis Jacq.) sometidas a ocho días en imbibición 1 (seis días en agua más dos días en BABA o en AS) antes de entrar a cuarto caliente, y a 20 días de calentamiento, presentaron en 82 días el mayor porcentaje medio de germinación óptima, alcanzando 34,44%, cuando se utilizó BABA, y 23,89%, cuando se usó AS, sin diferencia estadística entre los inductores. El tratamiento que presentó mayor porcentaje promedio (40%) de semillas germinadas con estándar óptimo fue el que permaneció en imbibición 1, seis días en agua más dos días en BABA, y 20 días de calentamiento. Por otra parte, el testigo tuvo un promedio de semillas germinadas óptimas de 1,11% en 150 días, presentando mayor incidencia de enfermedades (78,89%) con respecto a los factores (15,62%). Adicionalmente, con las dosis utilizadas de los inductores (2,5 mM BABA y 1 mM de AS), no se observaron daños o anormalidades en las semillas germinadas. Los hallazgos de esta investigación contribuirán a disminuir el tiempo de la germinación de las semillas de palma aceitera y a reducir los daños causados por patógenos durante el proceso de germinación.

