











 text in
text in  Spanish (pdf)
Spanish (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mail Cited by SciELO
Cited by SciELO  Cited by Google
Cited by Google  Similars in
SciELO
Similars in
SciELO  Similars in Google
Similars in Google 
 Permalink
Permalink
INTRODUCCIÓN
Los arrecifes de coral son ecosistemas altamente biodiversos y albergan las comunidades más ricas en especies de peces en el planeta (Sale, 2004). En términos generales los peces que habitan en los arrecifes coralinos se caracterizan por presentar un ciclo de vida con dos fases diferenciales, una primera fase larval pelágica que les permite dispersarse y una segunda fase adulta demersal relativamente sedentaria (Gerlach et al, 2007; Hamner et al, 2007; Hamner y Largier, 2012). Comprender la dinámica espacial y temporal de la fase larval es de gran interés debido a su importancia en el suministro de las poblaciones, los procesos de conectividad o intercambio de individuos entre las mismas y los patrones de dispersión de las especies (Cowen et al., 2000, 2006; Lefèvre y Bellwood, 2015). Las larvas de peces son quizás el grupo taxonómico mejor documentado dentro del plancton. Aunque en sus inicios la investigación en este grupo estuvo enfocada hacia la taxonomía, fisiología, etología, composición, abundancia y distribución, recientemente ha surgido un creciente interés en la comunidad científica marina por abordar problemas de estudio relacionadas con la dinámica poblacional de las comunidades ícticas, direccionado hacia conectividad y reclutamiento (Sale, 2004; Jones et al, 2005; Hogan et al,, 2012; Salles et al,, 2015) incluyendo estrategias y trayectorias (Irisson et al., 2004; Green et al., 2015).
En términos generales los estudios realizados sobre dispersión y distribución de larvas de peces apoyan la idea que desde los arrecifes de coral se exportan larvas hacia áreas cercanas, y posteriormente regresan para hacer un "auto-reclutamiento" de la población de la cual se originaron o bien pueden llegar a ser direccionadas a lugares cercanos (Harrison et al., 2012, Almany et al., 2013). Este tipo de información es relevante para determinar las zonas geográficas de importancia en el ciclo de vida de las especies que conforman una comunidad íctica particular, además de contribuir al conocimiento sobre el intercambio genético, la colonización de nuevos hábitats y el estado de las poblaciones (Buston et al., 2012).
Ha sido ampliamente documentado que los procesos que intervienen en el transporte de larvas y huevos funcionan a diferentes escalas espaciotemporales, destacándose los giros, el patrón superficial de circulación y los frentes térmicos o salinos entre otros. Además, factores biológicos como la tasa de depredación natural. Específicamente, estos elementos son responsables de la supervivencia, el transporte y el mantenimiento de las larvas en hábitats adecuados para su desarrollo o reclutamiento (Paris y Cowen, 2004; Hamner y Largier, 2012; Wolansky et al., 2014). En el Pacífico Oriental, las investigaciones realizadas sobre factores involucrados en los procesos de dispersión de larvas de peces han sido desarrolladas principalmente en Hawái, el Golfo de California y el Archipiélago Juan Fernández (Chile) (Bogan, 1994; Landaeta y Castro, 2004; Eble et al, 2011). Para el Pacífico de Colombia aún existen vacíos de información y escasez de conocimiento sobre la dinámica espacial y temporal del ictioplancton en ambientes costeros, a pesar de su importancia ecológica e interés económico y social que tienen las actividades pesqueras, siendo estos vacíos aún mayores para los ecosistemas de arrecifes de coral presentes en esta región del Pacífico Oriental Tropical.
Hasta la fecha, algunas investigaciones sobre las comunidades de peces adultos presentes en ambientes coralinos del Pacifico colombiano han sido desarrollados, principalmente en isla Gorgona (Rubio, 1990; Zapata y Morales, 1997; Zapata, 2001) y la ensenada de Utría (Estupiñan et al., 1990; Gómez y Viera, 1996). En estas investigaciones se ha estudiado principalmente la composición, abundancia y diversidad de este ensamble. Sin embargo, el conocimiento sobre el ictioplancton de estos lugares es reducido, aunque ha sido sugerido que la distribución espacial del zooplancton -incluyendo el ictioplancton- estaría dominado principalmente por el patrón de circulación local (Escarria et al., 2007; Giraldo et al., 2008, 2014; Giraldo y Valencia, 2012).
De acuerdo con Giraldo (2008), en isla Gorgona se identifican anualmente dos períodos oceanográficos contrastantes que modulan las condiciones ambientales del sistema pelágico, registrándose incluso cambios en la dirección prevaleciente de la circulación superficial que tiende a favorecer los procesos advectivos locales (Giraldo et al., 2008). Sin embargo, la presencia de la masa de isla sobre el campo de circulación superficial podría estar favoreciendo la formación de giros que podrían modular el patrón de distribución espacial de las larvas de peces como ha sido sugerido para otras localidades insulares (Boehlert et al., 1992; Cowen y Castro, 1994; Landaeta y Castro, 2004; Eble et al., 2011) favoreciendo la permanencia en el área de larvas de corta duración como son las larvas de las especies de peces que viven asociados a las formaciones coralinas (Searcy y Sponaugle, 2000; Zapata y Herron, 2002; Bergenius et al., 2005; Leahy et al, 2015). En la presente investigación se analizó la variación temporal y espacial del ensamble de estadios larvales de peces de especies, cuyo hábitat principal son las formaciones coralinas del sector oriental de isla Gorgona (lugar de mayor desarrollo de formaciones coralinas en la zona), considerando el efecto potencial de las condiciones oceanográficas locales sobre su composición y distribución espacial.
MATERIALES Y MÉTODOS
Área de Estudio
Gorgona es una isla continental ubicada en el océano Pacífico colombiano (2°58'10''N; 78°11'5''W) a 30 km del continente (Figura 1). La temperatura del aire en promedio es de 26 °C, mientras que la precipitación es de tipo bimodal con registros promedio anuales de 7000 mm, destacándose un periodo de baja precipitación de diciembre a marzo y dos momentos de máxima precipitación al año, uno en abril y el otro en noviembre (Blanco, 2009). La transparencia de la columna de agua varía de 6 a 12 m, la temperatura superficial del agua se encuentra entre 26 y 28 °C y la salinidad promedio mensual superficial oscila entre 28 y 33 unidades prácticas (Díaz et al., 2001; Giraldo, 2008; Giraldo et al., 2008).

Figura 1 Ubicación de la isla Gorgona y la malla de muestreo dentro de la zona de estudio. En gris localización de las formaciones coralinas en la isla Gorgona indicando AZ como arrecife La Azufrada, BL como arrecife Playa Blanca. Se indican los sectores definidos a priori con base en la distancia a las áreas de formaciones de arrecife de coral. S1= Sector 1. S2= Sector 2 y S3= Sector 3.
Para esta localidad han sido descritos dos periodos oceanográficos contrastantes: el primero, de mayo a diciembre con bajos valores de salinidad superficial y una termoclina profunda (entre 40 a 50 m de profundidad), y el segundo de enero a abril con alta salinidad superficial y termoclina somera (entre 5 a 10 m de profundidad). La marea es de tipo semidiurna, con registros máximos de altura de 5.7 m (Giraldo, 2008; Giraldo et al., 2008a). Esta variación intra anual de las condiciones oceanográficas locales en isla Gorgona está estrechamente relacionado con el efecto del desplazamiento latitudinal de la Zona de Convergencia Intertropical (ZCIT) sobre el Pacífico colombiano (Rodríguez et al., 2003; Blanco, 2009; Amador et al, 2016; Villegas et al., 2016). Específicamente, la ZCIT ha sido identificada como la principal fuente de variabilidad climática en la región (Poveda et al., 2011; Hoyos et al., 2013), además de modular el patrón de circulación oceánico y costero en el sector oriental del Pacífico oriental tropical incluyendo el Pacífico colombiano (Fiedler y Talley, 2006; Kessler, 2006; Villegas y Málikov, 2006; Rodríguez et al., 2007).
Las principales formaciones coralinas en el Pacífico colombiano se desarrollan sobre la franja oriental de isla Gorgona (Figura 1), destacándose los arrecifes de La Azufrada (AZ) con una extensión de 15 ha y el de Playa Blanca (PB) con una extensión de 9.6 Ha (Glynn y Ault, 2000). Al ser los arrecifes coralinos ecosistemas estratégicos, las formaciones coralinas de isla Gorgona han sido incluidas en el plan de manejo del Parque Nacional Natural Gorgona como objetos de conservación (UAESPNN, 2004; Muñoz y Zapata, 2013), siendo este uno de los principales temas de investigación en esta localidad durante los últimos 20 años (Giraldo et al, 2014).
Recolección de Muestras y Análisis
Para el desarrollo de esta investigación se estableció una malla de 16 estaciones oceanográficas en el sector oriental de isla Gorgona (Figura 1) y se seleccionaron dos períodos de muestreo: septiembre 2014 y febrero 2015, períodos que incluyen la variación intra anual de las condiciones oceanográficas que han sido descritas para esta localidad (Giraldo, 2008). En cada estación de muestreo se realizaron pescas horizontales superficiales (0-10 m) de zooplancton utilizando una red bongo de 30 cm de diámetro con flujómetros Hydrobios® adosados en cada boca y mallas colectoras de 250 um de poro. Previo a la pesca de zooplancton, se recolectaron muestras discretas de agua con una botella Niskin a 1 y 10 m de profundidad registrando la temperatura y salinidad utilizando una sonda multiparámetro YSIpro®. Además, se construyó el campo de circulación superficial instantáneo a 1 m de profundidad a partir del registro de la velocidad y la dirección del flujo en cada estación, mediante el seguimiento de un derivador pasivo tipo Davis modificado (Joseph, 2014; Jerez et al., 2017) que consistió en una boya inflable de 30 cm de diámetro, unido con una línea principal a un derivador de acero inoxidable con un peso muerto. El instrumento se dejó derivar por al menos 20 minutos en cada una de las estaciones, registrándose la coordenada geográfica inicial y final de deriva utilizando un GPS Garmin.
El zooplancton capturado fue conservado en una mezcla de formol y agua de mar al 4% buferizado con borato de sodio. En el laboratorio se separaron las larvas de peces y se realizó la identificación taxonómica hasta el menor nivel posible, utilizando las guías de ictioplancton de Moser (1996) y Beltrán y Ríos (2000). Para el desarrollo de los análisis, se seleccionaron las larvas de las especies de peces que han sido definidas por Rubio y Angulo (2003) y Robertson y Allen (2015) como especies asociadas a formaciones coralinas del Pacífico Oriental Tropical.
Análisis de datos
Se establecieron a priori tres sectores (S1, S2 y S3) en el área de estudio a partir de la distancia al arrecife coralino de La Azufrada (Figura 1). Se compararon las condiciones de temperatura. salinidad y abundancia de larvas de peces entre períodos utilizando una prueba no paramétrica de Mann-Whitney, y entre los sectores de estudio para cada periodo de muestreo utilizando una prueba no paramétrica de Kruskall-Wallis con corrección de Bonferoni y una prueba Post-hoc de rangos de Tukey.
La representatividad del muestreo se estableció como una proporción del valor esperado de riqueza para el área de estudio, definido a partir del estimador no paramétrico de riqueza Jackniffe de primer orden. Se comparó la riqueza de especies, dominancia y diversidad alfa entre periodos y entre sectores mediante un análisis de comparación por remuestreo de Bootstrap, a partir de 10000 interacciones aleatorias. Además, se estableció la tasa de recambio entre sectores a partir del índice de diversidad beta Morisita-Horn y se representó mediante un dendograma de similitud construido a partir de la función de ligamento promedio simple (Vellend, 2001; Jost et al., 2011; Calderón-Patrón et al., 2012), considerando el coeficiente de correlación cofenético como indicador de la significancia del agrupamiento establecido (Herrera-Moreno, 2000). Finalmente, se utilizó el método de interpolación inverse distance to a power grid del programa computacional Surfer11® para realizar la representación gráfica vectorial del campo de circulación superficial instantáneo y de la variación espacial de la abundancia de larvas de especies de peces asociados a formaciones coralinas en la zona de estudio.
RESULTADOS
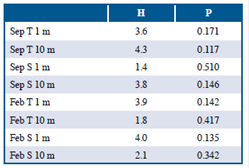
Las condiciones de temperatura y salinidad fueron contrastantes entre los periodos de muestreo en el área de estudio. Durante septiembre 2014, la temperatura superficial y a 10 m de profundidad fue significativamente mayor en comparación con febrero 2015 (Tabla 1). En contraste, durante febrero 2015 la salinidad superficial y a 10 m de profundidad fue significativamente mayor en comparación con septiembre 2014 (Tabla 1). Al evaluar la variación de estos parámetros oceanográficos entre los sectores de estudio, considerando cada período de muestreo de manera independiente, no se registraron diferencias significativas (Tabla 2). Este resultado sugiere que las condiciones térmicas y salinas fueron consistentes entre los sectores de estudios durante cada uno de los períodos de muestreo.
Tabla 1 Registros de temperatura y salinidad en la zona de estudio durante septiembre 2014 y febrero 2015. T = temperatura, S= salinidad, Z = valor del estadístico Z equivalente al estimador U de Mann-Whitney calculado, p = p valor, n = 9.

Tabla 2 Resultado del análisis de comparación de la temperatura (T) y salinidad (S) entre los sectores de estudio definidos durante cada periodo de muestreo. H = estimado Kruskall-Wallis, p = valor p.
El patrón superficial de circulación instantáneo durante septiembre exhibió dirección predominante hacia el oeste con una velocidad máxima de 21 cm s-1, mientras que durante febrero la dirección predominante fue hacia el noreste con una velocidad máxima de 52 cm s-1 (Figura 2). Estas condiciones hidrodinámicas en la zona de estudio sugieren que el patrón de circulación local podría estar influenciando la ubicación espacial de los organismos planctónicos, ya que durante septiembre la tendencia del desplazamiento por deriva sería hacia la zona costera de la isla mientras que durante febrero la tendencia sería hacia mar afuera (Figura 2).

Figura 2 Campo de circulación superficial instantáneo en el costado oriental de isla Gorgona durante septiembre 2014 y febrero 2015.
La representatividad del muestreo fue mayor a 74% en ambos períodos de estudio. En total fueron capturadas 1185 larvas (277 en septiembre y 908 en febrero) de 29 especies de peces asociados a formaciones coralinas en el área de estudio (Tabla 3). La abundancia, riqueza y diversidad del ensamble fue significativamente mayor en febrero que en septiembre (Tabla 4), estando dominado en septiembre por Coryphopterus urospilus (Gobiidae) y en febrero por C. urospilus y Stegastes sp. (Pomacentridae) (Tabla 3).
Tabla 3 Listado taxonómico de larvas de peces asociadas a arrecifes coralinos capturadas en el sector oriental de isla Gorgona. Se presentan los valores de abundancia relativa (Ab) y frecuencia de captura (Fr) durante los muestreos de septiembre 2014 y febrero 2015. Estaciones de muestreo por periodo: 16.

Tabla 4 Comparación de los atributos comunitarios entre septiembre 2014 y febrero 2015. Perm p: valor de significancia de la comparación por remuestreo de bootstrap a partir de 10,000 permutaciones.

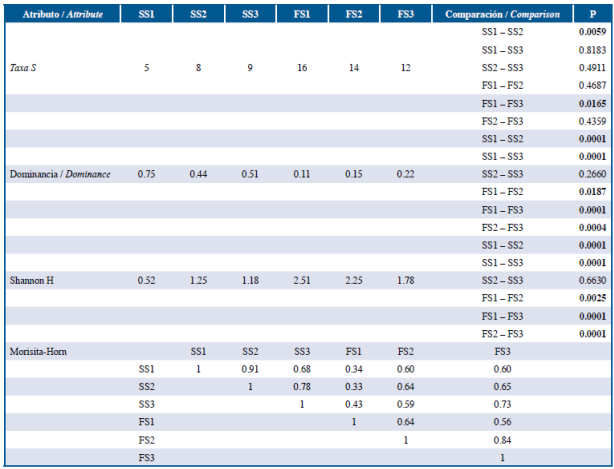
También se registraron diferencias significativas en los atributos comunitarios entre los sectores de estudio durante ambos periodos de muestreo (Tabla 5). Durante septiembre se registró un menor número de taxa en el sector más cercano a las formaciones coralinas, siendo en este sector significativamente mayor la dominancia y significativamente menor la diversidad y la riqueza de especies, mientras que durante febrero, la dominancia y la riqueza fueron significativamente menores en el sector más externo del área de estudio (Tabla 5).
Tabla 5 Atributos comunitarios del ensamble de larvas de peces asociados a formaciones coralinas en el sector oriental de isla Gorgona durante septiembre 2014 y febrero de 2015 considerando tres sectores durante cada periodo de estudio. SS1: septiembre sector 1. SS2: septiembre sector 2. SS3: septiembre sector 3. FS1: febrero sector 1. FS2: febrero sector 2. FS3: febrero sector 3 (diferencias significativas resaltadas en negrilla).
Algunas especies solo fueron registradas durante alguno de los periodos de estudio (Figura 3) ocasionando diferencias notorias en la composición de los ensambles, siendo la tasa de recambio entre periodos de 64% (Figura 3).
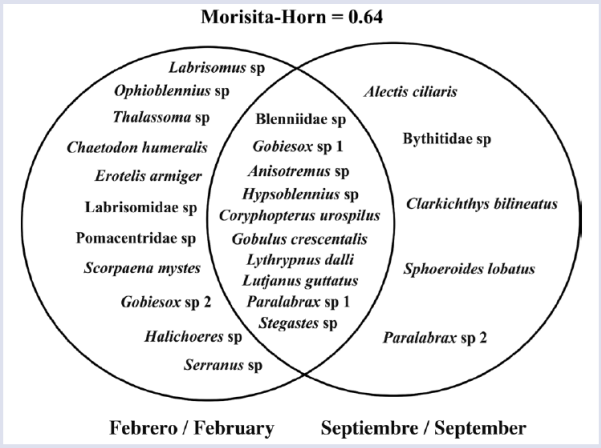
Figura 3 Descripción de la composición taxonómica de los ensambles de larvas de especies de peces asociados a formaciones coralinas en el área de estudio durante septiembre 2014 y febrero de 2015. Se indican las especies registradas exclusivamente en uno de los periodos y las especies compartidas (zona de traslape). Anisotremus sp. y Blenniidae sp. Contienen las especies nominales establecidas para cada uno de los géneros.
Este resultado sugiere variación estacional en los procesos reproductivos de las poblaciones de peces asociados a las formaciones coralinas en el área de estudio.
Al evaluar la variación de la diversidad del ensamble a lo largo del gradiente espacial definido (diversidad beta) se encontró que durante septiembre la mayor tasa de recambio de especie se presentó entre el sector más externo (S3) y el sector medio e interno, mientras que durante febrero la mayor tasa de recambio se presentó entre el sector interno (S1) y los sectores medio y externo (Figura 4, Tabla 5). Este resultado sugiere que el ensamble de larvas de especies de peces asociadas a formaciones coralinas en isla Gorgona no solo varía temporalmente, sino que también exhibió una variación espacial particular durante cada uno de los períodos de estudio.
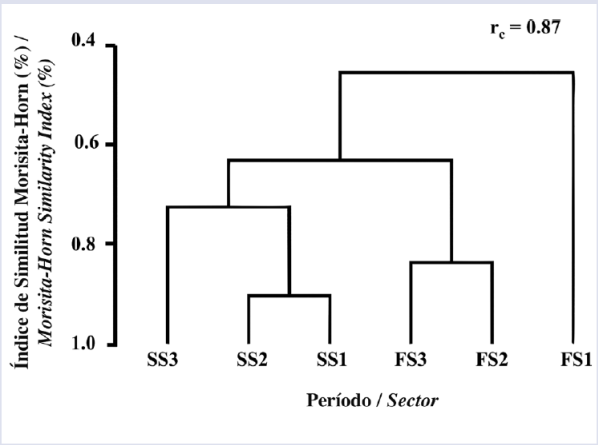
Figura 4 Dendograma de similitud del ensamble de larvas de especies de peces asociadas a formaciones coralinas por sectores de estudio en isla Gorgona, considerando los períodos septiembre 2014 y febrero 2015. rc: coeficiente de correlación cofenético.
La variación espacial de la abundancia total de larvas de especies de peces asociadas a formaciones coralinas fue diferente entre los períodos de muestreo (Figura 5). Durante septiembre, las mayores abundancias se registraron en el arrecife coralino de La Azufrada (1,925 larvas/1,000 m-3), conformando un centro de agregación (Figura 5). Durante febrero, los registros de abundancia en el sector del arrecife de La Azufrada fueron un orden de magnitud menor (68 larvas/1000 m3) a los de septiembre, identificándose durante este período una tendencia a incrementar la abundancia de larvas de especies de peces asociadas a formaciones coralinas en dirección noreste del área de estudio, específicamente hacia los sectores más alejados a las formaciones coralinas con registros del orden de 700 larvas/1,000 m3 (Figura 3).

DISCUSIÓN
Los ambientes insulares continentales y oceánicos han sido ampliamente reconocidos como entornos estratégicos para el desarrollo de las larvas de peces (Lobel y Robinson, 1986; Boehlert et al, 1992; Cowen y Castro, 1994; Paris y Cowen, 2004; Sale et al., 2005). Particularmente, isla Gorgona ha sido identificada como un área marina protegida estratégica para el Pacífico colombiano debido a su diversidad biológica y ecosistémica y su potencial función como sitio de abastecimiento de recursos pesqueros a nivel local (Franke y Acero, 1992; Franke y Acero, 1995a, 1995b; Rojas y Zapata, 2006; Giraldo et al, 2014).
Aunque el conocimiento sobre la biología y ecología de los estadios larvales de los peces es clave para entender la dinámica de sus poblaciones (Lehodey et al., 2006; Harrison et al., 2012), la información sobre este tópico en isla Gorgona es escasa. Recientemente, Escarria et al. (2007) registraron para isla Gorgona la presencia de larvas de 35 taxa de peces, de las cuales solo dos de ellas (Lythrypnus dalli y Paralabrax sp.) fueron registradas en la presente investigación. Por lo tanto, con los registros realizados durante nuestro estudio se incrementó el conteo de larvas de especies peces para isla Gorgona a 62 taxa.
Ha sido ampliamente documentado que la variación temporal de las condiciones ambientales puede llegar a modular la distribución, abundancia y, en general, la estructura de los ensambles de larvas de peces en el ambiente pelágico (Franco et al. 2002; Funes et al., 2002; Aceves et al., 2003, 2008). Durante el periodo de estudio, las condiciones oceanográficas registradas coincidieron con lo descrito previamente para esta localidad con una mayor temperatura y menor salinidad superficial durante septiembre (período de mayor precipitación) que durante febrero (periodo de menor precipitación) (Giraldo, 2008; Giraldo et al., 2008, 2012). Sumado a lo anterior, en la presente investigación se establecieron diferencias en la dirección predominante y velocidad media de la circulación superficial instantánea y diferencias en los atributos comunitarios de los ensambles de larvas de especies de peces asociados a formaciones coralinas entre los periodos de estudio. Por lo tanto, se puede inferir que las condiciones oceanográficas locales podrían estar condicionando la estructura y composición del ensamble de larvas de peces asociados a formaciones coralinas en esta localidad, ya sea modulando procesos reproductivos de los adultos o determinando la ubicación espacial de los huevos y larvas (Leis, 1993; Sponaugle et al., 2003; Fabricius et al., 2005; Srinivasan y Jones, 2006; Treml et al, 2012).
Es importante destacar que la abundancia de larvas de especies de peces asociadas a formaciones coralinas en el sector oriental de isla Gorgona también varió espacialmente dentro de cada uno de los períodos de estudio. Esta variación espacial "intraperíodo" podría estar relacionada con el patrón de circulación superficial, el cual durante septiembre estaría favoreciendo procesos de agregación hacía el sector donde se ubican las formaciones coralinas de La Azufrada y Playa Blanca mientras que durante febrero se estarían favoreciendo procesos de dispersión hacía toda el área de estudio. Estos resultados, concuerdan con lo descrito por Cowen y Castro (1994) a una escala de isla en Barbados, Antillas Menores. En esta localidad, la variación espacial de la abundancia de las larvas de peces asociadas a los arrecifes coralinos estuvo modulada por el campo local de circulación siendo las larvas transportadas activamente por la corriente, conformándose áreas de agregación. Incluso, ha sido propuesto que este sería un mecanismo utilizado frecuentemente por las larvas de peces coralinos para colonizar áreas aledañas o incluso retornar a los lugares de desove (Jones et al, 2005; Sundelöf y Jonsson, 2012).
En los estudios de dispersión, las larvas en sus primeros estadios de desarrollo son consideras como una partícula pasiva, cuya movilidad en los ambientes pelágicos insulares, estaría estrechamente relacionada con mecanismos físicos de la columna de agua, tales como corrientes, frentes termohalinos o remolinos de pequeña escala (Paris y Cowen, 2004; Hogan et al, 2012). Parece que la interacción de estos mecanismos físicos en una escala local, podrían llegar a favorecer la recirculación de las larvas de peces, reduciendo así la probabilidad de pérdida de individuos como consecuencia al transporte a lugares de asentamiento inadecuados ya que favorecerían el regreso de las larvas en la escala de días a los lugares de desove original (Jones et al., 2005, 2009; Hamner et al., 2007; Wolanski y Kingsford, 2014). Sin embargo, este mecanismo de permanencia puede verse aún más favorecido al considerar la capacidad sensorial (Lecchini et al., 2005; Mouritsen et al., 2013; Paris et al., 2013) y natatoria (Fisher et al., 2005; Irisson et al, 2015) que exhiben la mayoría de las larvas de peces coralinos, las cuales les permiten orientarse y dirigirse hacia lugares que les ofrezcan una mayor probabilidad de sobrevivencia.
Aunque los resultados de esta investigación pueden ser considerados como una visión sinóptica del área de estudio, se generó evidencia significativa que demuestra que en el sector oriental de isla Gorgona la disponibilidad de larvas de especies de peces asociados a formaciones coralinas varía temporal y espacialmente. Incluso, esta variabilidad parece estar relacionada con la variación temporal de las condiciones oceanográficas descritas para la zona de estudio, particularmente las condiciones de circulación superficial a escala de isla. Sin embargo, se hace necesario seguir ahondando en esta temática, incorporando en los estudios los factores que modulan la capacidad de movimiento de las larvas de peces, como su habilidad natatoria y sensitiva (Wolanski y Kingsford, 2014), de tal manera que se pueda llegar a comprender la forma como se estarían abasteciendo las poblaciones de peces que habitan las formaciones coralinas en esta localidad.

