











 Spanish (pdf)
Spanish (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mail Cited by SciELO
Cited by SciELO  Cited by Google
Cited by Google  Similars in
SciELO
Similars in
SciELO  Similars in Google
Similars in Google 
 Permalink
PermalinkINTRODUCCIÓN
Los crustáceos son uno de los grupos de mayor abundancia en los ambientes marinos, gracias a su gran capacidad de adaptación (ABELE, 1982; PONDER & LINDBERG, 2008). Ecológicamente juegan un papel importante en las comunidades macrobentónicas, debido a su abundancia y su posición intermedia dentro de las redes tróficas, siendo claves en la transferencia de energía hacia los niveles superiores como el de los peces demersales (FARIÑA et al., 1997; REDANT, 1982). Varias especies son objetivo de numerosas pesquerías en el mundo, siendo la pesca de camarón de aguas someras una de las más importantes (PAPACONSTANTINOU & KAPIRIS, 2003). En muchos casos la presión por obtener a corto plazo grandes cantidades de estos recursos han aumentado las tasas de captura (BOTSFORD et al., 1997; PAULY et al., 2003) llevándolos a la sobreexplotación (GUILLET, 2008). El Caribe colombiano no escapa a esta situación, lo que sumado con la falta de tecnificación y bajos ingresos obtenidos por la flota artesanal resume la problemática que enfrenta el sector pesquero en la región.
Como parte de las soluciones consideradas para dicha problemática se encuentra la pesca en aguas más profundas, lo que implica un gran reto teniendo en cuenta qué poco se conoce sobre la fauna de estos ambientes (CAMPOS et al., 2005; INVEMAR, 2010). En este sentido, estudios recientes referencian en el Caribe colombiano abundancias considerables de camarones como Aristaeomorpha foliacea, (Risso, 1827) (gamba española), Pleoticus robustus (Smith, 1885) (camarón rojo real), Penaeopsis serrata (Bate, 1881) (camarón rosado manchado) y Metanephrops binghami (Boone, 1927) (la langosta de aguas profundas), especies que presentan un alto valor comercial en varios mercados internacionales (BELCARI et al., 2003; PAPACONSTANTINOU & KAPIRIS, 2003) y que podrían ser recursos potenciales para la pesca de aguas profundas en el país (PARAMO & SAINT-PAUL, 2011a, 2011b, PARAMO & SAINT-PAUL, 2012).
Considerando lo anterior, se hace necesario profundizar en el conocimiento de los organismos de profundidad y el funcionamiento de estos ecosistemas; recolectar datos de abundancia y composición de especies y la relación con el ambiente que genere información de línea base. Por tal motivo, el objetivo del presente estudio fue determinar la composición y distribución de los crustáceos de aguas profundas capturados con nasas camaroneras en el área marina del departamento de Magdalena (Caribe colombiano).
METODOLOGÍA
El área de estudio comprendió el sector marítimo del departamento del Magdalena Caribe colombiano, desde la desembocadura del río Magdalena, hasta la desembocadura del río Palomino, siguiendo a INVEMAR (2000) en las ecorregiones Magdalena (MAG), Tayrona (TAY) y Palomino (PAL) (Figura 1). El diseño del crucero correspondió a un muestreo sistemático, con transectos paralelos y perpendiculares a la costa, espaciados regularmente cada 5 mn, desde 200 m hasta 500 m de profundidad, realizando muestreos cada 100 m. Las zonas de pesca se localizaron con una ecosonda científica Bisonic DT-X, con un transductor de 38 kHz y se registró su posición geográfica con un sistema de posicionamiento global (GPS por sus siglas en inglés) marca Garmin, modelo MAP 76CSx. Los muestreos fueron realizados entre octubre y noviembre de 2016, a bordo de un barco de pesca artesanal, usando nasas camaroneras plegables de 24"x 18" x 8", construidas con hilo de nylon multifilamento, un tamaño de malla de ½”, varillas de hierro de ½” y como carnada se utilizó machuelo entero y desmenuzado. Se construyeron tres trenes de 10 nasas cada uno, utilizándolos en cada estrato de profundidad, durante un periodo de 24 horas teniendo en cuenta las migraciones nictimerales de las especies. Los individuos capturados fueron identificados, pesados mediante una balanza de 0,1 g de precisión y medidos con calibrador con una precisión de 1 mm.

Figura 1 Área de estudio del departamento del Magdalena, Colombia, indicando las ecorregiones Magdalena (MAG), Tayrona (TAY) y Palomino (PAL).
Para evaluar si había diferencias significativas entre las abundancias entre las ecorregiones (en términos de número de individuos), se realizó la prueba no paramétrica de Kruskal-Wallis para muestras independientes. Se determinó la heterogeneidad de las especies de los crustáceos de profundidad calculando algunos de los principales índices ecológicos (diversidad de Shannon (H’), riqueza de Margalef (d), dominancia de Simpson (1-λ’) y equidad de Pielou (J’). Mediante el coeficiente de similitud de Bray Curtis, se estimaron las similitudes entre la composición de las especies para cada uno de los lances en términos de abundancia relativa (CLARKE & WARWICK, 1994); y se utilizó el análisis de escalamiento multidimensional (MDS) por medio del programa estadístico PRIMER v5.2.2 (CLARKE & GORLEY, 2001) para determinar grupos de similitud entre los puntos de muestreo.
RESULTADOS
Se realizaron 17 lances, siete en TAY, seis en PAL y cuatro en MAG, donde se capturaron 131 individuos en total. Taxonómicamente se identificaron 20 especies agrupadas en 12 familias, de las cuales seis son de interés comercial en otros países. A. spinicarpus, P. longipes y H. ensifer presentaron las mayores abundancias con 36, 24 y 23 individuos respectivamente, seguidas de las especies Myropsis quinquespinosa (12 individuos) y Plesionika edwarsii (siete individuos). Especies como E. bairdii, Plesionika tenuipes y Tetraxanthus rathbunae presentaron entre cuatro y cinco individuos y por último estuvieron especies como A. foliacea, Parapeneus politus y Plesionika sp. con un individuo cada una de ellas (Tabla 1). Se encontraron 14 especies en TAY, mientras que en PAL y MAG 10 y 8 respectivamente, sin embargo, no hubo diferencias significativas entre la abundancia en número de individuos por ecorregiones (N= 55; H= 3,618; 2 g.l.; valor p= 0,164).
Tabla 1 Especies de crustáceos decápodos encontrados durante los muestreos realizados entre octubre y noviembre de 2016, en las tres ecorregiones del área marina del departamento de Magdalena entre los 100 y 600 metros de profundidad. Con (*) se indican las especies con potencial interés comercial. Promedio=Promedio; Desviación estándar = d.e.; Valor máximo = Max; Valor mínimo = Min; especies con medida del ancho del caparazón (**).
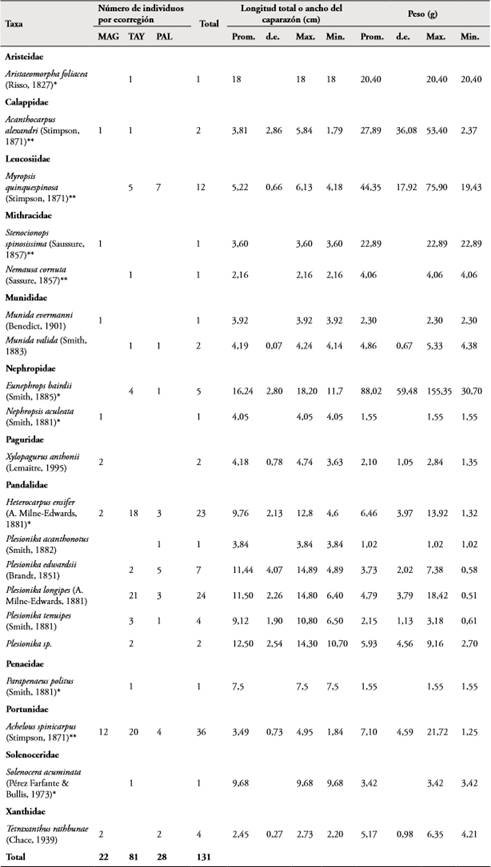
Las especies de mayor tamaño y peso fueron la langosta E. bairdii y el camarón A. foliacea, que alcanzaron tallas promedio de 18 cm y pesos de 88 y 20 gramos en promedio respectivamente. Las especies H. ensifer, P. longipes, P. tenuipes, Plesionika sp. y P. politus, son camarones de tallas medias que presentaron longitudes totales y pesos promedios entre los 8 y 14 cm y los 2 y 9 gramos, respectivamente. En cuanto a los cangrejos, M. quinquespinosa, Acanthocarpus alexandri y Stenocionops spinosissima fueron los de mayor tamaño en el ancho del caparazón y los más pesados (Tabla 1). Por lance, las especies que presentaron una mayor abundancia relativa en número de individuos fueron A. spinicarpus (2,57 ind/lance), P. longipes (1,71 ind/lance) y H. ensifer (1,64 ind/lance), mientras que en peso fueron M. quinquespinosa (0,038 kg/lance), E. bairdii (0,031 kg/lance) y A. spinicarpus (0,018 kg/lance). Este último cangrejo fue además el más frecuente (11 lances) seguido de los camarones H. ensiferus (siete lances) y P. longipes (seis lances) y el cangrejo M. quinquiespinosa (seis lances). Los individuos capturados se encontraron distribuidos en un rango de profundidad entre los 198 y 500 metros. Aunque A. spinicarpus, H. ensifer, Myropsis quinquespinosa y P. longipes presentaron amplios valores de distribución batimétrica, fueron más abundantes entre los 300 y 400 m (Figura 2).

Figura 2 Distribución batimétrica de los crustáceos capturados con nasas en el área marina del departamento de Magdalena, Caribe colombiano. Entre paréntesis se indica el total de individuos capturados de cada especie.
Los índices ecológicos mostraron mayor diversidad, equidad y predominio de crustáceos en PAL, a pesar de no tener la mayor riqueza de especies. TAY presentó la mayor riqueza de especies y número de individuos mientras que MAG obtuvo los menores valores en todos los índices (Tabla 2).
Tabla 2 Índices de diversidad por ecorregión para los crustáceos capturados con nasas en el área marina del departamento del Magdalena, Caribe colombiano. S: Número de especies, N: número de individuos, d: riqueza de Margalef, J’: equidad de Pielou, H’ (loge): diversidad de Shannon y 1-λ’: dominancia de Simpson.

En cuanto a la composición se encontraron tres agrupaciones, la primera conformada por lances realizados en TAY y PAL entre los 272 y 365 m de profundidad, caracterizados por tener un mayor número de especies y cantidad de individuos, siendo H. ensifer y A. spinicarpus las más comunes. La segunda agrupó lances de las tres ecorregiones entre los 280 y 417 m de profundidad con un número bajo de especies e individuos, siendo A. spinicarpus la especie dominante. La tercera agrupación correspondió a lances realizados en PAL y TAY entre los 198 y 364 m de profundidad, los cuales también presentaron un número bajo de especies e individuos y la especie dominante fue P. longipes (Figura 3).

DISCUSIÓN
Las bajas abundancias en número y peso de crustáceos obtenidas deben analizarse considerando que el esfuerzo de captura del presente estudio fue de un tren de 10 nasas operando 24 horas efectivas por lance por ser experimental, mientras que en una faena de pesca comercial se llegan a usar cuatro trenes de 290 nasas (GARCÍA-RODRÍGUEZ et al., 2000) y puede durar 30 horas (GÖNÜLAL et al., 2014). Aunque para el Caribe colombiano se han reportado altas biomasas de A. foliacea (mediante muestreos con pesca de arrastre) (PARAMO & SAINT-PAUL, 2011a), la baja abundancia de la especie en este estudio puede explicarse con las migraciones verticales relacionadas con la alimentación y la reproducción de ese camarón y con las que alcanza profundidades mayores a 600 m (D’ONGHIA et al., 1998; KAPIRIS & THESSALOU-LEGAKI, 2009).
Una de las especies de mayor abundancia en número de individuos y CPUE promedio fue el camarón P. longipes, coincidiendo con lo encontrado por PAJUELO et al. (2015) en el Atlántico centro-oriental (Islas Canarias), quienes encontraron que especies del género Plesionika dominan en la pesca con nasas en comparación con otros métodos de pesca como el arrastre. Otros autores afirman que la familia Pandalidae es muy abundante en aguas profundas del Mar Mediterráneo, el Mar Egeo, las Islas Canarias y Archipiélago de Madeira Atlántico nororiental, siendo Plesionika narval un recurso importante para la pesquería artesanal (ARCULEO et al., 2002; PAJUELO et al., 2015).
La mayoría de las especies se encontraron dentro del rango de distribución batimétrica reportado previamente por otros autores en el mundo (FELDER et al., 2009; HOLTHUIS, 1991; KING, 1984; KOMAI y KOMATSU, 2009; PEQUEGNAT & PEQUEGNAT, 1970; PÉREZ-FARFANTE & BULLIS 1973; WILLIAMS, 1984; POUPIN, 1994; RATHBUN, 1937) y para Colombia (CAMPOS et al., 2005; GÓMEZ-LEMOS et al., 2010; PARAMO et al., 2017), con excepción de P. politus que solo se había registrado hasta los 330 m (FELDER et al., 2009) y en el presente estudio se capturó por encima de los 400 m (Figura 2). La amplia distribución de profundidad de algunos crustáceos decápodos se debe a que mantienen un alto metabolismo a través de su comportamiento nectobentónico (COMPANY & SARDÀ, 1998) y son capaces de movilizarse a áreas de alimentación más favorables.
Coincidiendo con PARAMO et al. (2017), P. longipes fue la especie que mostró un mayor rango de profundidad (198 a 414 m, 280 m en promedio), lo que favoreció la mayor abundancia de este camarón en el área de estudio y por lo que podría ser considerada un recurso pesquero potencial. Sin embargo, debido a la poca información disponible sobre las especies y los ecosistemas de profundidad, y la incertidumbre sobre la biomasa existente, es necesario adoptar un enfoque precautorio hasta que se conozca la estructura y funcionamiento de dichos ecosistemas (FAO, 2003) y se profundice en la biología de esa especie en particular, así como prospecciones de biomasa y stock pesquero que permitan un aprovechamiento sostenible.
A. spinicarpus y M. quinquespinosa también fueron abundantes tanto en número de individuos como en biomasa, y aunque no se consideran comerciales están ampliamente distribuidas en el Caribe (WILLIAMS, 1984; RATHBUN, 1937). La importancia de estas especies crustáceos en los fondos marinos es que se encuentran dentro de un nivel trófico intermedio y son claves para el flujo de energía hacia los niveles superiores de la red (FARIÑA et al., 1997; REDANT, 1982). Por lo anterior, es de gran importancia tenerlas en cuenta en eventuales planes de pesquerías de profundidad para evitar alteraciones significativas dentro de los ecosistemas.
Las tallas encontradas para las especies de potencial importancia comercial se encuentran dentro de los ámbitos de tallas registrados para el Caribe colombiano (PARAMO et al., 2017). Para el caso específico de A. foliacea, el individuo encontrado superaba en tamaño la talla media de madurez reportada para el área (PARAMO & NÚÑEZ, 2015), situación que según los mismos autores, refleja la existencia de poblaciones sin explotar.
El suministro de alimentos es un factor esencial en la biodiversidad de aguas profundas (TECCHIO et al., 2011), la composición y estructura de estos ecosistemas son moduladas por la cantidad y calidad de alimento, ya sea por el enriquecimiento de la materia orgánica hundiéndose en la columna de agua (DANOVARO et al., 1999; GÖNÜLAL et al., 2014; SMITH et al., 2008) o por fenómenos de afloramiento (DANOVARO et al., 2003, GAGE 2003). Esta condición influye en la abundancia y la diversidad de la fauna bentónica (DANOVARO et al., 2008; LAMPADARIOU et al., 2009), la cual estuvo afectada, en el presente estudio, por las características intrínsecas de las ecorregiones, mostrando mayor similitud en las que se encuentran hacia la parte norte del área de estudio, donde la oceanografía local es modulada por eventos de surgencia estacional con alta productividad (GARCÍA-HOYOS, 2010; PARAMO et al., 2003; PARAMO et al., 2009). Esto concuerda con lo reportado en otros estudios donde la abundancia de camarones de profundidad está asociada con una alta productividad biológica (POLITOU et al., 2004), lo cual también podría explicar que en estas zonas se encontrara una mayor diversidad, sin embargo, con el número reducido de muestras los patrones de diversidad resultantes están lejos de ser definitivos.
Esta investigación provee información de gran importancia sobre la composición y distribución de los crustáceos de profundidad en el Caribe colombiano que son susceptibles a ser aprovechados, con métodos de pesca artesanal como las nasas, las cuales causan un menor impacto en el ecosistema que otros métodos habituales como la pesca de arrastre. Sin embargo, cabe considerar la realización de estudios que permitan determinar la biología y dinámica poblacional de estas especies potenciales para la pesca, generando bases para su manejo bajo un enfoque ecosistémico que garantice la sostenibilidad de las especies objetivo, la fauna acompañante y el ecosistema en el cual se encuentran.

