











 Spanish (pdf)
Spanish (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mail Cited by SciELO
Cited by SciELO  Cited by Google
Cited by Google  Similars in
SciELO
Similars in
SciELO  Similars in Google
Similars in Google 
 Permalink
PermalinkIntroducción
El orden Hemiptera está ampliamente distribuido en todo el mundo y se estiman 97.000 - 103.590 de especies conocidas, lo que representa alrededor del 7% de la diversidad de metazoarios (Zhang, 2011; Weirauchi y Schuh, 2011; Li et al., 2017). Este orden de insectos se divide en cuatro subórdenes: Auchenorrhyncha, Coleorrhyncha, Heteroptera y Sternorrhyncha (Cryan y Urban, 2012; Grazia et al., 2012). Sin embargo, las relaciones filogenéticas entre los grupos siguen siendo controvertidas y se ha cuestionado la monofilia de Auchenorrhyncha (Cryan & Urban, 2012; Song et al., 2012; Friedemann et al., 2014).
Los representantes de las 31 familias de Auchenorrhyncha (Urban & Cryan, 2007; Cryan y Urban 2012; Klimes et al., 2018) son terrestres y predominantemente fitófagos (Schuh & Slater, 1995). Las asociaciones ecológicas cercanas entre Auchenorrhyncha y sus hospedadores los convierten en indicadores ambientales, además de ser especialmente importantes como vectores de patógenos de plantas (Bartlett et al., 2018).
La familia Aethalionidae tiene solo tres especies con distribución en las regiones tropicales del mundo. Entre ellos, Aethalion reticulatum está ampliamente distribuido, y ocurre desde América Central hasta América del Sur (Marques, 1928; Barônio et al., 2012). Estos insectos son conocidos como cigarritas de árboles frutales, huertos o pedúnculos, A. reticulatum se alimenta de savia, lo que perjudica el desarrollo del fruto y el crecimiento de las plantas. Por lo tanto, las altas infestaciones de esta plaga pueden promover el retraso en el desarrollo y la caída de las frutas, por la succión continua de la savia, lo que puede causar grandes pérdidas agrícolas (Gallo et al., 2002; Santana et al., 2005; Henry, 2009; Oto et al., 2009; Rando & Lima, 2010; Camilo et al., 2013).
La morfología reproductiva genera caracteres que pueden, además de ayudar en la sistemática, contribuir al conocimiento del comportamiento reproductivo (Araújo et al., 2009, 2010, 2011, 2020; Özyurt et al., 2013; Dallai, 2014; Candan et al., 2018; Jiang et al., 2019; Munhoz et al., 2020) y en consecuencia para entender las explosiones demográficas y explorar técnicas de control (Parra, 2014).
En la mayoría de los estudios de morfología reproductiva de los Auchenorrhyncha se describen la ultraestructura del esperma (Folliot y Maillet, 1970; Cruz-Landim y Kitajima, 1972; Dai et al., 1996; Kubo-Irie et al., 2003; Chawanji et al., 2005, 2006; Dallai, 2014; SU et al., 2014; Dallai et al., 2016; Gottardo et al., 2016; Hodgson et al., 2016; Jiang y Qin, 2018; Jiang et al., 2019), destacando el estudio de la especie A. reticulatum (Araújo et al., 2010). Sin embargo, los estudios que describen la histología del tracto reproductivo en el suborden siguen siendo escasos (Robertson y Gibbs, 1937; Folliot y Maillet, 1970; Tian et al., 2006). Para contribuir al conocimiento de la familia Aethalionidae, se describe la morfología del sistema reproductivo de la A. reticulatum, teniendo como objetivo generar caracteres que puedan tenerse en cuenta en la sistemática del grupo, además de ampliar el conocimiento sobre la biología reproductiva de esta importante plaga agrícola.
Materiales y métodos
Los insectos. Se colectaron quince adultos machos de Aethalium reticulatum (Linnaeus, 1767) (L.) Blume (1856), de las ramas del arbusto Trema micrantha, en el Instituto de Biodiversidad y Sustentabilidad, (22°37’17” S y 41°78’57” W) de la Universidad Federal de Río de Janeiro, Macaé, Río de Janeiro, Brasil.
Microscopía de luz. Para el análisis histológico de los insectos, se crioanestesiaron y sus tractos reproductivos se diseccionaron en solución tampón de cacodilato de sodio 0,1 M, pH 7,2 y se transfirieron a un líquido Bouin durante tres horas a temperatura ambiente. Luego, las muestras se lavaron en tampón de cacodilato de sodio 0,1 M a pH 7,2, se deshidrataron en una serie cada vez mayor de concentraciones etanólicas (50, 70, 90 y 95%) y se sumergieron en historesina (Leica Historesin). Se obtuvieron secciones semifinas (2 µm de espesor) con una cuchilla de vidrio en un micrótomo de rotación (Leica RM 2255), posteriormente se coloreó con azul toluidina borato de sodio al 1% y se observó en un microscopio Olympus BX-50.
Resultados
El sistema reproductivo de los machos sexualmente maduros en A. reticulatum comprende un par de testículos, cada uno con cuatro folículos (Figura 1A), vesículas seminales, conductos deferentes, glándulas accesorias y un conducto eyaculador. En los testículos, cada folículo testicular se comunica en un conducto eferente y estos se fusionan para formar un conducto deferente (Figura 1A).
En cada folículo testicular, la espermatogénesis ocurre en diferentes zonas de desarrollo: zona de crecimiento (espermatogonias, donde ocurre la mitosis y se diferencia en espermatocitos, que se desarrollan sincrónicamente en quistes, formando haces o paquetes), zona de maduración (corresponde a dos divisiones meióticas que dan lugar a las espermátidas) y la zona de diferenciación (las espermátidas se alargan el flagelo y forman los espermatozoides) (Figura 1B). Los espermatozoides salen de los testículos en haces (Figura 1B). Dentro de cada folículo, los espermatozoides se desarrollan sincrónicamente en quistes mantenidos por la célula quística. En promedio en cada quiste se encontraron 649 espermatozoides (Figura 1C), en donde se muestran diez ciclos mitóticos durante la espermatogénesis.

Figura 1 Histología del sistema reproductivo de Aethalium reticulatum. A- Sección longitudinal del testículo que muestra los cuatro folículos testiculares (f). La flecha indica la región de unión de los conductos eferentes, que se unen para formar el conducto deferente. B- Folículo (f) que muestra las tres zonas de espermatogénesis: a = zona de crecimiento, b = zona de maduración, y c = zona de diferenciación. C- Quiste que contiene aproximadamente 649 espermatozoides. D- Conducto deferente pre-vesicular en la unión de los conductos eferentes. La flecha indica los núcleos de las células cúbicas. Barras = 200 µm.
Los conductos deferentes prevesiculares tienen un epitelio simple, formado por células cúbicas de núcleos esféricos y basales (Figura 1D). Los haces de los espermatozoides se deshacen cuando alcanzan las vesículas seminales (Figura 2A). La vesícula seminal es una región especializada que consiste en un engrosamiento en la región anterior del conducto deferente, en el cual los espermatozoides se almacenan hasta la cópula (Figuras 2A-B). La vesícula seminal presenta una forma ovalada (Figura 2A) y un epitelio simple formado por células cúbicas con núcleos esféricos (Figuras 2B-C). Una membrana basal separa las células epiteliales de una túnica formada por haces de células musculares (Figuras 2B-D). Los conductos deferentes postvesiculares tienen un epitelio simple formado por células cúbicas de núcleos esféricos y basales (Figura 2D). Las glándulas accesorias tienen una forma alargada (Figura 3A) y un epitelio formado por células prismáticas con núcleos ovales y muchos gránulos de secreción en todo el citoplasma y en el lumen de las glándulas (Figuras 3B-F).
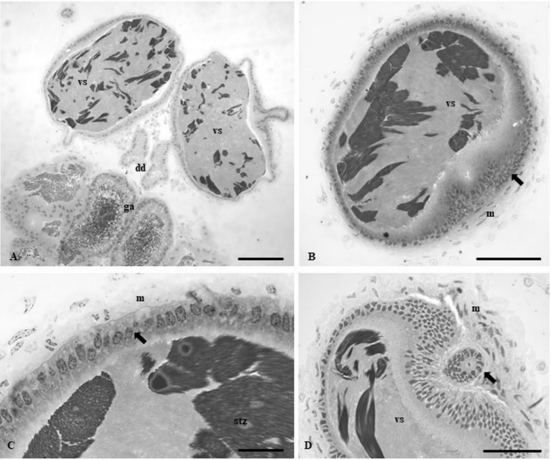
Figura 2 Histología del sistema reproductivo de Aethalium reticulatum. A- Sección longitudinal de la vesícula seminal (vs), conducto deferente (dd) y glándula accesoria (ga). B-C- Sección transversal de la vesícula seminal (vs), la flecha indica el epitelio cúbico con núcleos medianos, rodeado por una capa muscular (m). D- Vesícula seminal (vs) y región fr los conductos deferentes post vesiculares (flecha). Barras = 180 µm.

Figura 3 Histología del sistema reproductivo de Aethalium reticulatum. A- Sección longitudinal de las glándulas accesorias (ga). Las flechas indican la región de las glándulas accesorias (ga), con el lumen lleno de secreción. La flecha indica el epitelio. B-E- Se observa el detalle del epitelio de la glándula (flecha), que indica un epitelio prismático con núcleos apicales y el lumen lleno de secreción granular (vs) y región de los conductos deferentes postvesiculares (flecha). F- Región final de los conductos deferentes que conducen a la región de las glándulas. Las flechas indican núcleos basales en el epitelio de las glándulas, con varios gránulos de secreción en el citoplasma. Una capa muscular cubre todo el epitelio de los conductos deferentes. Barras = 150 µm.
Discusión
La morfología del sistema reproductor masculino de A. reticulatum sigue un patrón general de los insectos; con un par de testículos, conductos deferentes, vesículas seminales, glándulas accesorias y un conducto eyaculador, pero difiere de la mayoría de los Heteroptera (Adams, 2001; Lemos et al., 2005; Rodrigues et al., 2008; Freitas et al., 2010; Karakaya et al., 2012; Özyurt et al., 2013, 2014, 2015) por no tener un bulbo eyaculador.
En Hemiptera, en forma general se ha reportado que existe una gran variación en el número de folículos testiculares, que van desde dos en especies de Gerridae (Castanhole et al., 2008, 2010; Candan et al., 2018) y Miridae (Uceli et al., 2011) a siete folículos en varias familias de Heteroptera (Pendergrast, 1957; Kumar, 1965; Karakaya, 2012; Gomes et al., 2013; Kaur y Patia, 2012, 2016; Freitas et al., 2007, 2010; Özyurt et al., 2014; 2015). En A. reticulatum se observaron cuatro folículos, sin embargo, no es posible establecer una comparación dentro del suborden Auchenorrhyncha debido al bajo número de taxones previamente estudiados.
Los folículos en diferentes zonas de desarrollo en machos maduros demuestran que estos insectos continúan produciendo espermatozoides durante la edad adulta. La producción continua de espermatozoides es común en especies que tienen una larga vida adulta y copulan a lo largo de esta fase (Buschini, 2007; Moreira et al., 2008) y este patrón espermatogénico se describe para muchas otras especies de Heteroptera (Jamieson et al., 1999; Pires et al., 2007; Rodrigues et al., 2008).
La formación de haces de espermatozoides en los testículos y su desenlace ocurre cuando alcanzan las vesículas seminales, esto también se encontró en otros insectos (Quicke et al., 1992; Moreira et al., 2004; Lino-Neto et al., 2008). El número de espermatozoides en los haces o paquetes representan el número final de células que se desarrollaron en sincronización dentro de los quistes, a partir de una única espermatogonoia (Lino-Neto et al., 2008). A. reticulatum muestra 10 ciclos mitóticos, en himenópteros, el número de ciclos mitóticos ha sido una característica señalada para su uso en análisis sistemáticos (Araújo et al., 2010; 2012), sin embargo, para Hemiptera en general, aún no es posible agregar este carácter en las inferencias filogenéticas.
El epitelio que se encuentra en los conductos espermáticos a lo largo del tracto reproductivo es similar al descrito para otros insectos como Heteroptera (Karakaya, 2012; Gomes et al., 2013; Kaur y Patia, 2012, 2016; Freitas et al., 2007, 2010; Özyurt et al., 2014; 2015), en abejas sociales (Dallacqua & Cruz-Ladim, 2003; Araújo et al., 2005; Lima et al., 2006; Brito et al., 2010), hormigas (Ball y Vison, 1984; Wheeler y Krutzsch, 1992) y avispas (Dirks y Sternburg, 1972; Bushrow et al., 2006; Moreira et al., 2008). Sin embargo, se pueden ver algunas diferencias en los epitelios de las glándulas accesorias y sus diferentes tipos de secreción. Las secreciones de las glándulas accesorias están asociadas con el mantenimiento y la activación de los espermatozoides y la inducción y aceleración de la oviposición en las hembras (Chen, 1984; Raina et al., 1994; Gillot, 2003). En A. reticulatum estas glándulas están bien desarrolladas y en los machos sexualmente maduros están llenas de secreción. Es posible que tales secreciones estén involucradas en las estrategias reproductivas del macho, relacionadas con mecanismos que pueden garantizar la fidelidad de la cópula y mantener sus espermatozoides viables en la espermateca de la hembra durante un período más largo.
El conducto eyaculatorio, como en la mayoría de los insectos, es único, mediano y tiene una cutícula que muestra su origen ectodérmico (Bushrow et al., 2006; Moreira et al., 2008). Los espermatozoides de A. reticulatum ya han sido descritos por Araújo et al., (2010), generando caracteres importantes para futuras inferencias en el sistema Auchenorrchyncha.
En este trabajo describimos la histología del sistema reproductivo de A. reticulatum, ampliando el conocimiento sobre la estrategia reproductiva de este insecto, que puede usarse para generar mecanismos para controlar esta plaga agrícola.

