











 English (pdf)
English (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mail Cited by SciELO
Cited by SciELO  Cited by Google
Cited by Google  Similars in
SciELO
Similars in
SciELO  Similars in Google
Similars in Google 
 Permalink
Permalink
INTRODUCTION
Changes in fish diet in Neotropical currents may be related to seasonal and environmental variations (Abelha et al. 2001), such as the presence or absence of riparian vegetation, aquatic vegetation, and the availability of resources in the environment that can vary drastically in Andean streams between hydrological periods. For this reason, aspects such as understanding the trophic ecology of the fish that live in these streams, the way in which they use resources due to migration patterns among different streams and rivers during different stages of their life, as well as the changes in diet occurring over time by variations in the availability of prey between hydrological periods are important to support the conservation measures and management actions for species that, such as the B. henni, are subject to strong pressures due to deterioration of riparian banks, overfishing and high pollution levels.
Brycon henni (Eigenmann, 1913) known as sabaleta in Colombia, spreads only in the Magdalena-Cauca River basin (Montoya-López et al. 2006; Mancera-Rodríguez, 2017), and in the study zone it is part of the subsistence fishing resource of the inhabitants of nearby municipalities. This species is not listed in the risk categories in the red book of freshwater fishes from Colombia (Mojica et al. 2012), but was subsequently assessed and classified in the category of least concern (LC) in the Red List of Threatened Species of the International Union for Conservation of Nature - IUCN (Villa-Navarro et al. 2016). Sabaleta is a fish with omnivorous behavior (Flórez, 1999; Magallanes & Tabares, 1999; Botero-Botero & Ramírez-Castro, 2011), and short lateral migrations with routes of less than 100 km have been documented among the main courses of the rivers and adjacent streams for reproductive purposes (Martínez-Orozco & Vásquez-Zapata 2001; Arboleda-Chacón et al. 2005).
Uncontrolled fishing, the impact on water bodies related to the change in land use that has generated deforestation due to livestock activities and intensive use of chemicals in agriculture, as well as pollution due to urban wastewater settled in the basins of Nare and Guatapé rivers have drastically reduced the population sizes of this species (Mancera-Rodríguez, 2017).
The species spawns in streams and individuals that are born remain there and grow until they reach a size close to that of sexual maturity, when they move to the main river bed where their adult stage goes on. Therefore, it is important to study the changes in diet associated with these movements during the development of the species and the effect of the change in vegetation cover on the banks of the water bodies where it lives. Especially because for omnivorous species, such as the sabaleta, food resources in the form of leaves, seeds, fruit and insects are mainly supplied by riparian vegetation (Esteves & Aranha, 1999). This is why the presence of forest cover in the banks of streams and rivers is important for their foraging behavior (Vitule et al. 2008; Abilhoa et al. 2010).
Therefore, the objectives of this study were: (i) to characterize the trophic ecology of sabaleta Brycon henni in tributary streams of Nare and Guatapé rivers, in the Magdalena River basin, Colombia, and (ii) to analyze the variations between sexes, size class, different degrees of anthropic intervention and among hydrological periods (dry, dry-to-rainy transition and rainy periods). Understanding the trophic ecology features of the species will help establish recommendations for its protection, conservation, and management.
MATERIALS AND METHODS
Area of study. The work was carried out in eight stations located on tributaries of Nare and Guatapé rivers in the municipalities of Concepción, San Roque, Alejandría, San Rafael and San Carlos, department of Antioquia, in the Magdalena River basin, Colombia (Table 1). Six stations were established in the Nare River basin in streams with high anthropogenic disturbance, without tree cover on their banks, and with land uses dedicated to agricultural and livestock activities. Two stations were set in the Guatapé River basin in water bodies that had riparian vegetation without alterations or with a low degree of anthropogenic disturbance. The characteristics of the water bodies studied, their habitats and their physical-chemical characteristics are detailed in Mancera-Rodríguez (2017).
Sampling. The specimens were collected in five samplings carried out in the rainy period (October and November 2007, and May 2008), dry-to-rainy transition period (March 2008), and in dry period (January 2010). The specimens were landed with SAMUS 725MP electro-fishing and 1 cm mesh eye nets. They were labeled and preserved in 70 % alcohol in the Ecology and Wildlife Conservation laboratory of the Universidad Nacional de Colombia, Sede Medellín.
Analysis of stomach contents. A total of 262 stomachs of B. henni individuals were examined, of which 147 were landed in the rainy period, 64 during the dry-to-rainy transition, and 51 in the dry period. Of these, 119 were females, 127 males, and 16 immature specimens whose sex could not be determined (Table 1). For each specimen, its standard length SL (mm) was measured with a digital caliper, and its total weight (g), eviscerated weight (g) and weight of each stomach contents were obtained with an analytical balance (Ohaus Scout-Pro 202; 0.01 g). The specimens were eviscerated for the extraction of the digestive tracts, which were weighed (g) and preserved in 70 % alcohol. Subsequently, the separation of the food categories was carried out with an EZ4 stereoscope (Leica Microsystems). The emptiness index (EI) was calculated as EI = (Se/St) x 100, where Se is the number of empty stomachs and St the total number of stomachs analyzed.
Table 1 Location of sampling stations and number of individuals of sabaleta (Brycon henni) landed in the Nare and Guatapé river basins, Magdalena River system, Colombia.
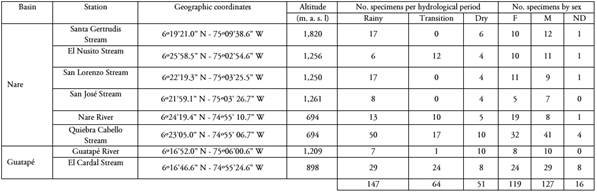
F: Females M: Males ND: Not determined
A dietary category accumulation curve was used to assess the sufficiency of sample size of B. henni stomachs containing identifiable prey for all data sets. The mean and standard deviation of the cumulative number of new preys were calculated and the sufficiency of the sample size was statistically evaluated using the linear regression method of Bizzarro et al. (2007).
Dietary components were separated and identified qualitatively and quantitatively according to volumetric and frequency of occurrence methods (Ricker, 1971; Hyslop, 1980), and the Alimentary Importance Index (AII) according to Kawakami & Vazzoler (1980) AII = Fi*VI /
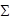 (Fi*Vi), where AII = alimentary importance index, Fi = frequency of occurrence (%) of each category and Vi = volume (%) of each category.
(Fi*Vi), where AII = alimentary importance index, Fi = frequency of occurrence (%) of each category and Vi = volume (%) of each category.
Feeding categories were identified at the lowest possible taxonomic level and AII data were analyzed using a non-parametric Kruskall-Wallis analysis of variance (Zar, 2010) to detect differences in consumption rates (AII) for each food category by sex, size classes, hydrological period and between the basins of Nare (high anthropogenic alteration) and Guatapé (low anthropogenic alteration) rivers. Post hoc comparisons were also made and evaluated through the Mann-Whitney test, using PAST software (Hammer et al. 2001). To evaluate size-related variations in feeding habits, specimens were separated into size classes of 20 mm standard length, taking into account the interval estimation made by Mancera-Rodríguez (2017) in the study of reproductive aspects of the species in these two basins of Nare and Guatapé rivers.
Clustering analysis was performed on standardized AII values using the Bray-Curtis similarity index (based on abundance) using the paired grouping method to describe variations by size classes of feeding habits using PAST software (Hammer et al. 2001).
The trophic niche breadth for the utilization of food resources was calculated by size, to define whether there are marked changes in dietary specialization, using AII values according to Levin's Niche Amplitude measure (Krebs, 1999): Bi =
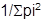 , where pi is the proportion with which each category of item i contributes to the diet. To standardize this measure of niche breadth to a 0 to 1 scale, the standardized niche breadth was calculated: B = Bi-1/n-1 where, B is the Levin’s standardized niche breadth, Bi is the Levin’s niche measure and n is the number of items in the diet. The B value is close to zero when individuals have minimal diet breadth or maximum specialization.
, where pi is the proportion with which each category of item i contributes to the diet. To standardize this measure of niche breadth to a 0 to 1 scale, the standardized niche breadth was calculated: B = Bi-1/n-1 where, B is the Levin’s standardized niche breadth, Bi is the Levin’s niche measure and n is the number of items in the diet. The B value is close to zero when individuals have minimal diet breadth or maximum specialization.
The Morisita’s Diet Overlapping Index (Krebs, 1999) was calculated for each hydrological period to define the diet overlapping between groups I and II. The Morisita’s simplified index is shown in equation 1:
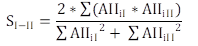 equation 1
equation 1where AIIiI is the Alimentary Importance Index of the ith food in group I and AIIiII is the Alimentary Importance Index of the same item i in group II. Diet overlapping increases as Morisita’s index increases from 0 to 1. Values close to 1 indicate high dietary overlapping where individuals compete for the same dietary components.
RESULTS AND DISCUSSION
Characteristics of the specimens and the diet. The size range of the sampled specimens varied from 42.7 to 241.0 mm SL and the weight between 1.2 and 288.1 g. The mean standard length (SL) was 121.2 mm, and the mean total weight was 45.5 g. The average size for females evaluated was 127.9 mm SL (53.5-241.0) and for males 120.2 mm SL (60.0-232.5). By basin, individuals from the Nare River basin had an average size of 118 mm SL (53.5-234.0) and for the Guatapé River basin 127.3 mm SL (42.7-241.0). No significant differences were detected in the size of the specimens evaluated or between basins, sexes or hydrological periods evaluated (Mann-Whitney, p>0.05).
The diet category accumulation curve to evaluate the sufficiency of the number of samples reached a plateau above 242 stomachs analyzed, which provides a good description of B. henni diet. Of the total number of stomachs evaluated, only one was empty, with an emptiness index of 0.38 %. Stomach content was classified into 20 types of prey that were grouped into seven food categories (Table 2). Two categories were classified as primary: plant material made up of seeds, fruit, and algae (AII: 59.65 %), and the Insecta class made up of aquatic macroinvertebrates of various orders and remains of non-identifiable insects (AII: 37.80 %) (Table 2). The other five categories were considered secondary with little contribution to the diet: fish, made up of fish remains and scales (AII: 1.95 %), Nematoda (AII: 0.58 %), and the Crustacea, Arachnida and Mollusca categories with an AII less than 0.01 %, each (Table 2).
Table 2 Composition of the sabaleta (Brycon henni) diet in the Nare and Guatapé river basin, as a percentage of frequency of occurrence (%F), percentage of Volume (%V) and percentage of the Alimentary Importance Index (AII) for all sampled stomachs (n=262).
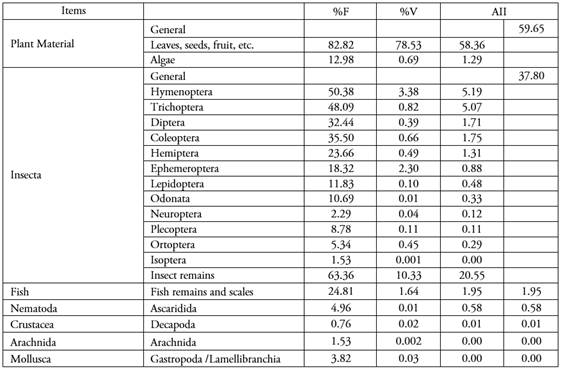
In the middle basins of the Nare and Guatapé rivers, B. henni feeds mainly on plant material and can be considered a herbivorous/frugivorous species (fruit, stems, leaves, roots, as well as various types of grass and legume seeds), which complements their diet with insects (both aquatic larval phases and allochthonous origin) mainly of the orders Trichoptera, Hymenoptera, Diptera, Coleoptera, Hemiptera, and Ephemeroptera, and to a lesser extent other groups such as fish, crustaceans and arachnids. In this regard, it has been pointed out that several species of the Brycon genus are consumers and seed dispersers of various species of riparian plants (Horn, 1997; Gomiero et al. 2008; Reys et al. 2009; Correa et al. 2015).
The species has a wide range of prey consumed and has been considered an omnivorous species, but its diet is made up mainly of plant material, and to a lesser extent prey of animal origin, including several orders of insects, fish remains, crustaceans, mollusks, annelids and even frogs (Flórez, 1999; Magallanes & Tabares, 1999). However, Botero-Botero & Ramírez-Castro (2011) and Feriz García et al. (2023) found that the species had a generalist diet with characteristics of a carnivorous species, with a tendency to consume mainly aquatic insects and non-native organisms, and to a lesser extent to consume plant material in the Portugal de Piedras river (Valle del Cauca), and in the Timbío river (Cauca), respectively. In that sense, Restrepo-Santamaría et al. (2022) found that the species fed on a wide variety of terrestrial and aquatic invertebrates, as well as plant material and seeds for the Porce III reservoir in the department of Antioquia.
The above shows that the sabaleta is a fish species that exhibits opportunistic eating habits and easily adapts to the different environments and aquatic environments of the Colombian Andes (Jiménez-Segura et al. 2016), which has allowed it to adapt even to the lentic environments generated by the construction of reservoirs in various water courses where the species is spread (Martínez-Orozco & Vásquez-Zapata, 2001; Restrepo-Santamaría et al. 2022).
Changes in diet by size and effect of anthropogenic intervention. Diet composition by sex did not have significant differences in terms of the AII of the primary food categories in general. Therefore, the following analyses were performed by grouping the two sexes. The frequency of the main prey groups in the B. henni diet by size class shows plant material and insects as the primary food source across size categories (Figure 1). However, specimens with an SL greater than 170 mm had a higher percentage of plant material and a lower percentage of insects in the AII and specimens with a SL greater than 210 mm had an increase in the share of fish (Figure 1).
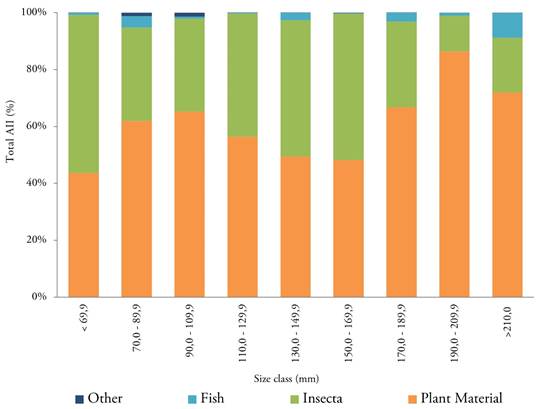
Figure 1 Frequency of the primary prey groups in the sabaleta (Brycon henni) diet by size classes (standard length) in the Nare and Guatapé river basins, Magdalena River system, Colombia. AII: Alimentary Importance Index.
Grouping analysis using the Bray-Curtis similarity index based on AII values for the 9 size classes distinguished two main groups: group I (fish smaller than 170.0 mm SL) and group II (fish larger than 170.0 mm SL) (Figure 2).
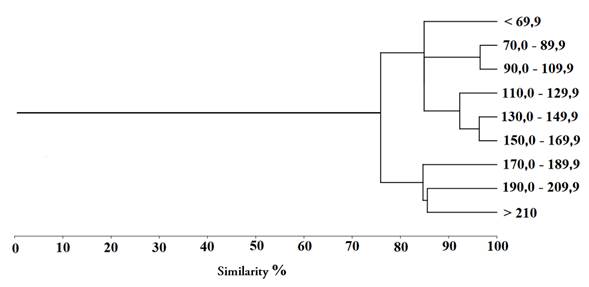
Figure 2 Bray-Curtis similarity grouping dendogram based on Alimentary Importance Index (AII) values by size classes for sabaleta (Brycon henni) in the Nare and Guatapé river basin, Magdalena River system, Colombia.
Significant differences were found between the two size groups in fish consumption that was higher for group II (Mann-Whitney, P=0.041). Despite the values not being significant, a higher consumption of plant material and a lower consumption of insects were found in individuals larger than 170 mm SL (Table 3).
Insect consumption was important in specimens smaller than 170 mm SL and higher in streams with greater anthropogenic impact and less tree cover on their banks. Specimens with an SL greater than 170 mm showed higher consumption of plant material and less insect consumption, and specimens with an SL greater than 210 mm showed an increase in fish amount in their diet. These ontogenic changes are consistent with the reports about other species of the same genus by Drewe et al. (2004), who found that B. guatemalensis went from a carnivorous diet of insects in younger fish to a diet of fruits and leaves as adults in streams in Costa Rica. Similar results were found by Albrecht et al. (2009) for B. gouldingi and B. falcatus in Brazil, who pointed out that smaller individuals mainly consumed terrestrial arthropods and larger individuals mainly consumed seeds and fruit.
Between basins with different degrees of anthropogenic intervention, differences in diet were observed with greater consumption of plant material (Mann-Whitney, P=0.009) and fish (Mann-Whitney, P=0.021) in the Guatapé River basin that presents low anthropogenic alteration, and greater consumption of insects in the Nare River basin that has a higher degree of anthropogenic intervention (Mann-Whitney, P=0.004) (Table 3). This is related with the findings by other authors who have associated the presence of forest cover on the riparian banks of the streams with a greater consumption of leaves, seeds, fruits, but also insects that are mainly supplied by riparian vegetation (Esteves & Aranha, 1999; Vitule et al. 2008; Abilhoa et al. 2010). It is also consistent with the reports for species of the Brycon genus that largely consume non-native foods, showing the importance of maintaining riparian forests (Gomiero et al. 2008; Albrecht et al. 2009; Reys et al. 2009).
Table 3 Variation in the Alimentary Importance Index (AII) of the different categories of prey in the sabaleta (Brycon henni) diet by sex, size groups and basin in the Nare and Guatapé rivers, Magdalena River system, Colombia. SL = Standard Length
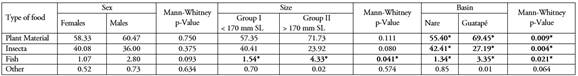
* Statistically significant differences (p<0.05)
However, the significant increase in insect consumption in the streams of the Nare River basin, whose banks have undergone high deterioration and loss of forest cover, shows that those species that take advantage of non-native foods, such as the sabaleta can modify their feeding habits due to changes in riparian vegetation.
Variation in diet for different hydrological periods. During the rainy period, a greater consumption of plant material was observed in the sabaleta specimens evaluated in the two size groups and there were no significant differences (Mann-Whitney, p>0.05) in the consumption of the different food categories. However, during the dry period, Group I had differences with a lower consumption of plant material (Mann-Whitney, P=0.031) and fish (Mann-Whitney, P=0.001) compared to Group II and a higher consumption of insects in its diet (Mann-Whitney, P=0.041). Additionally, during the transition period from dry season to rainy season, Group I had differences with a lower consumption of plant material (Mann-Whitney, P=0.010) compared to Group II and a higher consumption of insects (Mann-Whitney, P=0.010) (Table 4). Levin's Niche Breadth was lower in the rainy period (B=0.080 and 0.131 for Group I and II, respectively) because the diet was based primarily on plant material for the two groups. During the dry period it was B=0.262 for Group I and B=0.255 for Group II, and in the transition period B=0.242 for Group I and B= 0.300 for Group II, with a tendency towards a more generalist diet in these two periods. Similarly, Correa & Winemiller (2014) found that Brycon falcatus and B. melanopterus exhibited frugivorous feeding habits during the rainy season and their diet changed to insectivorous habits in the dry season in the Colombian Amazon region.
Table 4 Variation of the alimentary importance index (AII) of the different categories of prey in the sabaleta (Brycon henni) diet by size groups in the different hydrological periods in the Nare and Guatapé river basins, Magdalena River system, Colombia. Group I: < 170 mm SL, Group II: > 170 mm SL.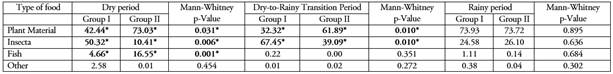
High overlapping in the species diet was detected between both size groups with a Morisita’s index value of S(I-II) = 0.957. Diet overlapping had its maximum value during the rainy period (S(I-II) = 0.995), in which the two groups fed mainly on plant material and insects, mostly on the orders Hymenoptera, Trichoptera, Diptera, Coleoptera and Hemiptera. In the transition period the value was S(I-II) = 0.833 with a greater consumption of insects by individuals smaller than 170 mm SL and a decrease in consumption of plant material. In the dry season, the Morisita’s index value was the lowest (S(I-II) = 0.803), because Group I had a lower consumption of plant and fish material, and a higher consumption of insects than Group II. For the three periods, the value indicates a strong overlapping in the diet for the species with consumption of the same food groups, although with different proportions in each group depending on the hydrological period.
Frugivorous species such as B. henni have been strongly affected by fishing, deforestation from livestock activities and pollution due to the use of chemicals in agriculture on the banks of streams in the Nare and Guatapé river basins. This has caused a decrease in size and in average size at sexual maturity (Mancera-Rodríguez 2017), which affects their potential as possible seed dispersers, which is carried out by larger fish.

