










Services on Demand
Journal
Article
 Spanish (pdf)
Spanish (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mailIndicators
-
 Cited by SciELO
Cited by SciELO -
 Access statistics
Access statistics
Related links
-
 Cited by Google
Cited by Google -
 Similars in
SciELO
Similars in
SciELO -
 Similars in Google
Similars in Google
Share

 Permalink
PermalinkRevista Facultad Nacional de Agronomía Medellín
Print version ISSN 0304-2847
Rev. Fac. Nac. Agron. Medellín vol.58 no.2 Medellín July/Dec. 2005
MASA TOTAL Y CONTENIDO DE NUTRIENTES EN RAÍCES FINAS DE ECOSISTEMAS FORESTALES (Pinus patula Schltdl y Cham Cupressus lusitanica Mill y Quercus humboldtii Bonpl.) DE PIEDRAS BLANCAS, ANTIOQUIA-COLOMBIA
TOTAL FINE ROOT MASS AND NUTRIENT CONTENT IN FOREST ECOSYSTEMS (Pinus patula Schltdl and Cham Cupressus lusitanica Mill and Quercus humboldtii Bonpl.) FROM PIEDRAS BLANCAS, ANTIOQUIA-COLOMBIA
Luis Hernán Barreto Sánchez1 y Juan Diego León Peláez2
1 Ingeniero Forestal. Universidad Nacional de Colombia, Sede Medellín. Facultad de Ciencias Agropecuarias. A.A. 1779, Medellín, Colombia. <lhbarret@unalmed.edu.co>
2 Profesor Asociado. Universidad Nacional de Colombia, Sede Medellín. Facultad de Ciencias Agropecuarias. A.A. 1779, Medellín, Colombia. <jdleon@unalmed.edu.co>
Recibido: Junio 28 de 2005; aceptado: Octubre 10 de 2005.
RESUMEN
En la región de Piedras Blancas, Antioquia - Colombia, se realizó un muestreo de raíces finas (£ 5 mm) en tres parcelas permanentes establecidas en bosques de pino pátula (Pinus patula Schltdl y Cham), ciprés (Cupressus lusitanica Mill) y roble (Quercus humboldtii Bonpl.) a diferentes profundidades: 0-30, 30-50 y 50-80 cm, con el fin de determinar la masa de raíces, y la concentración y contenido de nutrientes en cada cobertura vegetal. La masa de raíces finas disminuyó rápidamente a medida que aumentaba la profundidad, encontrándose en la parcela de ciprés el 83 % de las raíces en los primeros 30 cm de profundidad, en pino pátula el 75 % y en roble el 68 %. Los elementos con mayor participación en las raíces finas fueron calcio y potasio, siendo el primero de ellos mucho mayor en la cobertura de ciprés con respecto a las parcelas restantes, seguido en abundancia del potasio, a excepción de la parcela de roble en donde la concentración de un oligoelemento como Fe, fue incluso superior a la de calcio. Los elementos Ca, Mg, K, P, Mn, Cu y Zn acumulados en las raíces finas, hasta los 80 cm de profundidad, fueron superiores en la cobertura de ciprés, con la única excepción de Fe, que fue superior en la parcela de roble. En las parcelas de pino pátula y ciprés, el patrón de abundancia siguió la secuencia decreciente Ca > K Mg > Fe > P. En este trabajo se pretendió resaltar la importancia de las raíces finas y la concentración de los elementos, en los ecosistemas forestales naturales y plantados y su relación con el ciclo de nutrientes. Para ello, la aproximación metodológica comprendió la masa total de raíces, esto es raíces vivas y muertas en conjunto.
Palabras clave: Pinus patula, Cupressus lusitanica, Quercus humboldtii, raíces finas, ciclo de nutrientes.
ABSTRACT
In the Piedras Blancas region, Antioquia-Colombia, a sampling of fine roots (£ 5 mm) was conducted in three permanent plots established in forest ecosystems of pine (Pinus patula Schltdl), cypress (Cupressus lusitanica Mill) and oak (Quercus humboldtii Bonpl.), at different depths: 0-30, 30-50 and 50-80 cm, in order to determine the mass of roots and the concentration and content of nutrients in each vegetation type. The mass of fine roots diminished quickly as depth increased, representing in the cypress plot 83 % of all roots in the first 30 cm of depth, in pine 75 %, and in oak forest 68 %. The elements with the highest participation in the fine roots were calcium and potassium, with the former much greater in the cypress cover compared to the other plots, continuing in abundance potassium, with the exception of the oak plot where the concentration of Fe microelement was even greater than that of calcium. The elements Ca, Mg, K, P, Mn, Cu, and Zn accumulated in the fine roots up to 80 cm of depth were greater in the cypress cover, with the only exception being Fe, which was greater in the oak plot. In pine and cypress plots, the pattern of abundances followed the following sequence Ca > K > Mg > Fe > P. This study sought to highlight the importance of fine roots and their concentrations of nutrients in natural forests and plantations and their relationship with nutrient cycling. For this purpose, the methodological approach comprised total root mass, both dead and alive.
Key Words: Pinus patula, Cupressus lusitanica, Quercus humboldtii, fine roots, nutrient cycles.
 MATERIALES Y MÉTODOS
MATERIALES Y MÉTODOS
RESULTADOS
DISCUSIÓN
CONCLUSIONES Y RECOMENDACIONES
RECONOCIMIENTOS
BIBLIOGRAFÍA
Distintos estudios han demostrado la importancia de las raíces finas para el funcionamiento del ecosistema, no sólo por su participación en la productividad primaria neta (PPN), sino también en los ciclos biogeoquímicos de los bosques, debido a que su descomposición contribuye significativamente al enriquecimiento de los horizontes orgánicos por su rápida reconversión (Burke y Raynal, 1994). Los estudios de biomasa de raíces finas en los trópicos son mucho más abundantes que los de raíces gruesas, debido a su importancia en la adquisición de recursos para la vegetación e igualmente como fuente de materia orgánica del suelo (Sanford y Cuevas, 1996).
Las raíces finas se distribuyen principalmente en los primeros centímetros del suelo y disminuyen exponencialmente a medida que aumenta la profundidad. Se ha estimado que en bosques húmedos tropicales el 57 % de la biomasa de raíces finas se encuentra en los primeros 30 cm del suelo (Jackson Money y Schulze, 1997). Sin embargo, la profundidad hasta la cual pueden penetrar las raíces finas está relacionada con la tasa de infiltración de nutrientes y las relaciones hídricas hasta 30-40 cm de profundidad. Además, la distribución horizontal de las raíces finas está altamente correlacionada con la concentración de nutrientes intercambiables del suelo, especialmente fósforo y calcio. En sitios donde hay una alta disponibilidad de nutrientes se localiza una alta densidad de raíces finas y micorrizas (Stark y Spratt, 1977).
Por otra parte, el crecimiento de las raíces finas es modificado por una compleja interacción entre las raíces y cambios en los factores ambientales, en el tipo de actividades o por la presencia de hormonas que regulan el crecimiento. No obstante, el potencial de interacción de numerosos factores afecta el crecimiento de las raíces, la temperatura o la combinación temperatura y precipitación pueden ser usados para explicar la variación observada en la masa y volumen de raíces finas. En muchos casos los factores climáticos y la latitud se correlacionan con el volumen de raíces finas, pero muy pobremente con la masa de raíces finas vivas. Esto sugiere que el clima tiene más fuerte influencia sobre el volumen de raíces finas que sobre la masa viva de raíces finas (Vogt et al., 1997).
La biomasa de raíces, por sí sola, no es un buen indicador de la capacidad de absorción de recursos por las plantas, la densidad de las raíces es un mejor indicador, ya que relaciona la longitud radical por unidad de volumen de suelo. Sin embargo, es claro que las raíces finas son las encargadas de la captura de los recursos en la planta, entre mayor sea su biomasa, mayor será la capacidad adquisitiva (Vogt et al., 1997). Además, la biomasa de raíces finas está directamente relacionada con condiciones favorables para la fotosíntesis, la estructura y composición florística del bosque, la disponibilidad de nutrientes y su variación con micrositios, con la presencia de oxígeno y con la porosidad y permeabilidad del suelo. Sin embargo, en el estudio de la biomasa de raíces finas muchos autores no han prestado atención a la separación entre vivas y muertas, estimando así la masa de raíces finas, compuesta por la biomasa y la necromasa (Klinge y Herrera, 1978).
En este trabajo se pretende resaltar la importancia de las raíces finas y la concentración de los elementos, en los ecosistemas forestales naturales y plantados y su relación con el ciclo de nutrientes. Para ello, la aproximación metodológica comprende la masa total de raíces, esto es raíces vivas y muertas en conjunto.
Área de estudio. La zona de estudio se localiza al oriente del departamento de Antioquia - Colombia, Cuenca hidrográfica de la quebrada Piedras Blancas (Figura 1). Geográficamente está ubicada en las coordenadas planas del sistema nacional cartográfico (Plancha IGAC 147-I-D) Y: 840.500 – 841.500, X: 1.188.000 – 1.189.000; su altitud aproximada es de 2.490 m.

Figura 1. Localización del área de estudio empleada para cuantificar la masa total y contenido de nutrientes en raíces finas. Piedras Blancas, Antioquia-Colombia
La temperatura media anual es de 14,9 ºC, la humedad relativa promedio anual es de 82 %, y la precipitación media anual es de 1.698 mm. El régimen climático es isotérmico. La precipitación es bimodal, concentrándose los mayores valores en dos períodos: marzo-mayo y septiembre-noviembre. Las condiciones climáticas señaladas corresponden con la zona de vida (Holdridge, 1987) bosque húmedo Montano Bajo (bh-MB).
En la Cuenca aproximadamente el 35 % del área son plantaciones forestales (1.020 ha) establecidas en su mayoría en la década de los 60, de las cuales pino pátula (Pinus patula) representa cerca de 330 ha y ciprés (Cupressus lusitanica) 570 ha. Dentro de los bosques naturales (1.270 ha en la cuenca), se destaca como formación ecológica el ecosistema de robledal (Quercus humboldtii) (Empresas Públicas de Medellín (EPM), 1989). Dichas coberturas vegetales, en la zona de estudio, presentan profundidades de la capa de hojarasca que oscilan entre: 35-40 cm en pátula, 5-10 cm en ciprés y 40-60 cm en roble.
Trabajo de campo. El trabajo se realizó en las parcelas experimentales establecidas desde el año 2001 donde se han monitoreado diferentes aspectos de los ciclos de agua y nutrientes. Tales parcelas han sido dispuestas en bosques de pino pátula (Pinus patula), ciprés (Cupressus lusitanical) y roble (Quercus humboldtii), y cada una de ellas con una extensión de 5000 m2. Algunas de las características de estos bosques se muestran en la Tabla 1.
Tabla 1. Caracterización de los rodales utilizados para el estudio de la masa total y contenido de nutrientes en raíces finas. Piedras Blancas, Antioquia-Colombia.

Para la estimación de raíces finas (diámetro< 5 mm) se establecieron seis puntos de muestreo según arreglo sistemático en cada una de las parcelas. Para la extracción de cada muestra se utilizó en campo un barreno de martillo tipo Slide Hammer AMS, el cual consta de un cilindro de 35 cm de largo por 4,7 cm de diámetro interno, conectado por su parte superior con un martillo que permite golpear verticalmente para la penetración del cilindro en el suelo. Antes de introducir el barreno, era retirada la capa de hojarasca con la ayuda de palas pequeñas y escobillas; posteriormente se extrajeron las muestras de suelo diferenciadas en tres niveles de profundidad: nivel cero de 0- 30 cm, nivel uno de 30- 50 cm y nivel dos de 50- 80 cm de profundidad. El material se almacenó en bolsas de polietileno con sello hermético debidamente marcadas y se colocaron en neveras de icopor de forma que conservaran la temperatura normal y se transportaron al laboratorio. En total se extrajeron 54 muestras.
Trabajo de laboratorio. El material obtenido en campo se almacenó en un cuarto de enfriamiento -Cava del Laboratorio de Ecología y Conservación Ambiental (LECA) de la Universidad Nacional de Colombia, Sede Medellín- a una temperatura de 4 °C. De allí fueron retiradas para su procesamiento, siendo las muestras lavadas con un elutriador (Fahey et al., 1999), el cual permite disolver la muestra de suelo y separar el material vegetal con la ayuda de agua a presión.
En este trabajo se utilizó el sistema de lavado construido para la investigación Producción y descomposición de raíces finas en bosques tropicales maduros y jóvenes de la región de Porce, Antioquia (Sierra, 2004; Usme, 2004), el cual fue especialmente adaptado del modelo presentado por Martyn y Ross (1996). Este sistema consta de una cámara inferior en forma de campana, una torre de 60 cm de altura en tubo de PVC de 5 y una línea horizontal igualmente en tubo de PVC de 3 que posee en su parte final una malla de tamiz de 0,54 mm de diámetro. La cámara a su vez tiene conectados dos inyectores de agua a presión a ambos lados del extremo inferior.
La muestra de suelo se coloca en esta cámara y el agua se hace circular a una velocidad moderada y constante, por lo que el movimiento ascendente hace que las partículas más pequeñas (limo y arcilla) salgan por la descarga y pasen a través de los orificios del tamiz, mientras que las partículas de arena permanecen en la base del equipo, las raíces, los residuos orgánicos y algo de arena fina quedan retenidos en el tamiz. La circulación de agua por el lavador se suspende cuando el agua de la descarga luce clara y presumiblemente todas las raíces han sido extraídas del suelo. Posteriormente, todo el material recogido en el tamiz era colocado en agua y la arena fina eliminada mediante procesos repetidos de lavado. Estas muestras eran luego empacadas en pequeños trozos de tela debidamente marcados, y llevadas al laboratorio para su conservación en refrigerador a baja temperatura, hasta su procesamiento.
Finalmente, las muestras fueron separadas manualmente con la ayuda de pinzas metálicas, se depositaron en bandejas de aluminio y se llevaron al horno a 80 °C durante 24 horas para hallar su peso seco. Debe destacarse que la separación realizada no diferenció entre raíces finas vivas y raíces finas muertas -objetivo por fuera de los alcances de este trabajo- siendo por tanto los valores de masa de raíces finas mencionados a lo largo del artículo, los correspondientes a la masa total que comprende la suma de ambas fracciones. Estas muestras fueron llevadas posteriormente al laboratorio para determinación del contenido de nutrientes. Para las determinaciones analíticas, debido a que para este tipo de análisis se necesita un peso considerable de raíces finas, fue necesario combinar las muestras obtenidas de las tres profundidades en cada cobertura, con lo cual se obtuvo una concentración promedia para cada una de ellas por elemento. Los elementos determinados fueron Ca, Mg, K, P, Cu, Fe, Mn y Zn, mediante espectrofotometría de absorción atómica y ultravioleta visible (P). Todos los análisis y procesamiento de muestras se realizaron en el LECA. Para efectos de comparación y análisis de estos resultados con los niveles de fertilidad del suelo en las distintas parcelas, fueron empleados los análisis practicados en ellas por León (2005).
Análisis estadístico. Previa realización de los análisis de varianza (ANAVA), fueron practicadas pruebas de normalidad a los datos mediante el método de Shapiro-Wilks. Para la comparación de la masa de raíces finas entre cada una de las coberturas y entre profundidades, se realizó un ANAVA para un diseño factorial de tipo 3 x 3. Finalmente, fueron empleadas pruebas de rangos múltiples (método de Tukey HSD con un nivel de significancia del 95 %) para la determinación de diferencias estadísticamente significativas entre medias. Para todos los análisis fue empleado el programa estadístico STATGRAPHICS PLUS 5.0.
Masa de raíces finas. De conformidad con los resultados presentados en la Tabla 2, se puede afirmar que los datos se distribuyen normalmente (P<0,01). El ANAVA indica que los factores cobertura y profundidad tienen un efecto estadísticamente significativo sobre el peso de raíces finas (P < 0,05), así como las interacciones entre estos dos factores.
Tabla 2. Análisis de varianza para peso de raíces finas en ecosistemas forestales de Piedras Blancas, Antioquia-Colombia.

La masa de raíces finas (diámetro < 5 mm) disminuye muy rápidamente con la profundidad (Figura 2), se observa claramente que el peso de raíces finas es muy superior en los primeros 30 cm de suelo. En la parcela de pino pátula aproximadamente el 75 % del peso total de raíces finas (6,07 mg ha-1) se encuentra en los primeros 30 cm de suelo, en la cobertura de ciprés es de aproximadamente 83 % (14,75 mg ha-1) y en la cobertura de roble es de aproximadamente el 68 % del peso total (4,23 mg ha-1).
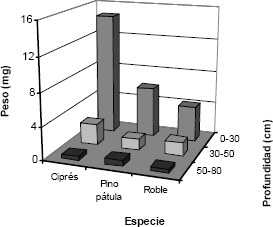
Figura 2. Peso de raíces finas (mg ha-1) en las tres especies forestales determinado a tres profundidades, en la región de Piedras Blancas, Antioquia-Colombia.
Las Tablas 3, 4 y 5 muestran los resultados de la prueba de Rangos Múltiples de medias entre coberturas para las profundidades 0-30, 30-50 y 50-80 cm, respectivamente. Para la primera profundidad -0-30 cm- (Tabla 2) se observa que las coberturas pino pátula y roble, y las coberturas pino pátula y ciprés, forman grupos homogéneos. Además, se puede afirmar con un nivel de confianza del 95 % que existe diferencia estadísticamente significativa entre los pesos de raíces finas de las coberturas ciprés y roble para esta profundidad.
Tabla 3. Prueba de Rango Múltiple para el peso de raíces finas por cobertura forestal a la profundidad 0- 30 cm, en la región de Piedras Blancas, Antioquia-Colombia.

Tabla 4. Prueba de Rango Múltiple para el peso de raíces finas por cobertura forestal a la profundidad 30- 50 cm, en la región de Piedras Blancas, Antioquia-Colombia.

Tabla 5. Prueba de Rango Múltiple para el peso de raíces finas por cobertura forestal a la profundidad 50- 80 cm, en la región de Piedras Blancas, Antioquia-Colombia.

Para las profundidades 30- 50 cm y 50- 80 cm (Tablas 4 y 5) se observa que las tres coberturas forman un grupo homogéneo. Además, se puede afirmar con un nivel de confianza del 95 % que no existe diferencia estadísticamente significativa entre los pesos de raíces finas de las tres coberturas para ambas profundidades.
Nutrientes en las raíces finas. En la Tabla 6 se muestran los valores medios de concentración de elementos presentes en las raíces finas –vivas y muertas- de las parcelas de pino pátula, ciprés y roble, los cuales sirvieron de base para calcular las cantidades totales almacenadas de cada elemento. En la Tabla 7 se presenta el capital de nutrientes acumulado en las raíces finas para los tres niveles de profundidad, así como los pesos medios de raíces finas en las tres parcelas.
Tabla 6. Concentración de elementos en las raíces finas de tres especies forestales establecidas en la región de Piedras Blancas, Antioquia-Colombia.
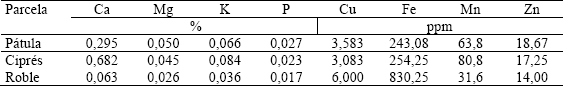
Tabla 7. Peso promedio de raíces finas y elementos acumulados a tres profundidades en especies forestales establecidas en la región de Piedras Blancas, Antioquia-Colombia.

Como patrón general puede señalarse que los elementos con mayor participación en las raíces finas son calcio y potasio. La concentración del primero es mucho mayor en la cobertura de ciprés con respecto a las parcelas restantes, mientras que el potasio le sigue en abundancia, siendo la única excepción la parcela de roble, en donde la concentración de un oligoelemento como Fe, es incluso superior a la de dicho macronutriente. Con relación a Fe, cabe asimismo resaltar que alcanza concentraciones incluso por encima de P en las parcelas de ciprés y roble, y mayores que Mg en la parcela de roble (Figura 3).

Figura 3. Concentración de macroelementos (a) y microelementos (b) en las raíces finas de tres especies forestales establecidas en la región de Piedras Blancas, Antioquia-Colombia.
Se encontró que los elementos Ca, Mg, K, P, Mn, Cu y Zn acumulados en las raíces finas, hasta los 80 cm de profundidad, fueron superiores en la cobertura de ciprés (Figura 4), con la única excepción del oligoelemento Fe, que fue superior en la parcela de roble. En las parcelas de pino pátula y ciprés, el patrón de abundancia siguió la secuencia decreciente Ca > K > Mg > Fe > P, siendo muy similares los valores de estos dos últimos elementos en la parcela de pino pátula. Como excepción del patrón descrito, se tiene que en la parcela de roble, el elemento mayoritario es Fe, por encima incluso de calcio, conforme asimismo ocurrió con las concentraciones para dicho elemento en esta misma parcela.

Figura 4. Contenido total de nutrientes en las raíces finas de tres especies forestales establecidas en la región de Piedras Blancas, Antioquia-Colombia.
.
Masa de raíces finas. La proporción de raíces finas en la parte superior del suelo – 0-30 cm – fue bastante alta: 75 %, 83 % y 68 % bajo las coberturas de pino pátula, ciprés y roble, respectivamente. Esto evidencia que la masa de raíces finas disminuye muy rápidamente con la profundidad (Lundgren, 1978; Burke y Raynal, 1994; Wassenaar, 1995; Moreno-Chacón y Lusk, 2004).
La cantidad de raíces finas en los primeros 30 cm de suelo siguió la secuencia decreciente ciprés > pino pátula > roble, lo cual coincide con los resultados obtenidos por Lundgren (1978) en Tanzania, según los cuales la cantidad de raíces finas en plantaciones de ciprés, en diferentes sitios, era mayor que aquella presente en rodales de pino pátula. En el caso particular de dicho estudio, las mayores cantidades de raíces finas se correspondían, para ambas especies, con unos menores niveles de fertilidad edáfica, lo cual sugeriría la compensación de la baja disponibilidad de nutrientes con una mayor capacidad de exploración del suelo para su captura, lograda a través de una masa de raíces más abundante. En las parcelas, sin embargo, tal asociación no resultó clara, ya que los niveles de fertilidad a esta profundidad, son muy semejantes para ambas plantaciones.
Cabe destacar sin embargo, que cuando se compara la masa de raíces objeto de estudio, determinada por Lundgren (1978) para los primeros 30 cm del suelo con la de este trabajo, y se tiene en cuenta la disponibilidad de nutrientes, se verifica claramente la asociación entre el nivel de fertilidad del suelo y la cantidad de raíces. Así, mientras para Lundgren (1978), los parámetros de fertilidad edáfica eran claramente superiores a los de este estudio, las cantidades de raíces en las parcelas de Piedras Blancas, son por contrapartida visiblemente mayores para ambas plantaciones. De esta forma, la masa total de raíces –vivas y muertas- en la parcela de ciprés fue de 2,8 a 5,7 veces superior a la encontrada en los rodales de esta especie en Tanzania y, en el caso de pino pátula, de 1,6 a 2,9 veces las correspondientes al estudio en mención.
Un gran número de hipótesis han sido propuestas alrededor del desarrollo de la masa de raíces. La infertilidad extrema de muchos de los suelos por debajo de ellas y la efectividad que muestran para la retención de nutrientes ha llevado a su interpretación como producto de la selección natural para la retención de nutrientes (Stark y Jordan, 1978; Kingsbury y Kellman, 1997). En este sentido, se ha encontrado que la concentración de raíces finas en la superficie del suelo y en la hojarasca, están frecuentemente asociados con los ciclos cerrados de nutrientes, como respuesta a subsuelos infértiles y dependencia del reciclaje biológico. Sin embargo, hay desventajas en tales distribuciones, así, en el caso particular de la captura de agua para las plantas, excesivas concentraciones de las raíces en cercanías a la superficie no resultan beneficiosas (Baillie, 1996).
Se ha mostrado además que una característica de los bosques húmedos tropicales, es que estos suelen presentar una alta cantidad de raíces finas en la capa superficial del suelo, aunque esto puede variar de acuerdo al tipo de suelo, al tipo de bosque y a las especies que allí se encuentren (Sanford y Cuevas, 1996). Sin embargo, de conformidad con Vogt, Grier y Vogt (1986) la masa de raíces finas más alta medida en los trópicos fue reportada por Cuevas y Medina (1983) para bosques húmedos no caducifolios de Venezuela, cuyos suelos se caracterizan por presentar una alta acumulación de materia orgánica profunda y una baja disponibilidad de nutrientes.
Como regla general, Wassenaar (1995) menciona que -con base en la revisión de varios estudios- el desarrollo de raíces es superficial en un gran número de sitios observándose una relación directa entre el desarrollo de tales raíces y la disponibilidad de nutrientes en el mantillo, en el agua lluvia, en las aguas de escorrentía por el tallo, y en el agua que gotea desde las copas de los árboles.
Se han sugerido que la proliferación de raíces finas se encuentra estimulada por la materia orgánica muerta (St. John, 1983). En este sentido, como se desprende de la Tabla 7, la masa de raíces en la parcela de roble en los primeros 30 cm del suelo, es notoriamente inferior a aquella presente en las parcelas de coníferas. Sin embargo, la cantidad de raíces presente en la hojarasca acumulada sobre el piso forestal –mantillo- es muy superior en esta parcela, contabilizando allí 3,91 mg ha-1, 0,96 mg ha-1 en la parcela de pino pátula y 1,65 mg ha-1 en la parcela de ciprés (León, 2005). Tal situación sugiere el desarrollo de mecanismos de conservación de nutrientes por la formación natural, llegando a invadir incluso el material recientemente desprendido del dosel arbóreo y acumulado sobre el piso del bosque.
A este respecto, de conformidad con los hallazgos de Herrera et al. (1978) y Cuevas y Medina (1988), existe evidencia que soporta incluso la captura directa de algunos nutrientes por las raíces desde el material acumulado sobre el piso y sometido al proceso de descomposición, siendo éste el caso de K, Ca, Mg y P. Adicionalmente, la presencia en estos ecosistemas de hongos micorrizales asociados a las raíces finas, aseguran la captura y el almacenamiento de los nutrientes disponibles. Estos dos mecanismos, operando en conjunto, permiten el mantenimiento de un ciclo cerrado de nutrientes (Ross, 1993).
Una hipótesis alternativa explica la cantidad de raíces como una respuesta a ambientes del suelo mineral que resultan inhóspitos para aquellas, caso de la ocurrencia de estaciones de anegamiento, y altas concentraciones de Al intercambiable (Stark y Spratt, 1977; St. John, 1983).
Como un posible factor que incidiría en una alta concentración de raíces en las capas más superficiales del suelo, se tiene la verificación de unos bajos niveles de fertilidad edáfica en los horizontes más profundos. Sin embargo, esta situación no corresponde con la encontrada en este estudio, ya que la mayor fertilidad para la segunda profundidad de muestreo -30-50 cm- ocurre precisamente en la parcela de ciprés, en donde se tienen las mayores cantidades de raíces finas en los primeros 30 cm del suelo. Al igual que con el primer nivel de muestreo, para el segundo se encuentra correspondencia entre la disponibilidad de nutrientes y la masa de raíces en el caso de la parcela de ciprés; en las parcelas de roble y pátula, en donde los niveles de fertilidad son muy similares, se presentan así mismo cantidades semejantes de raíces finas.
Dado el rango tan amplio definido por los valores de masa de raíces finas de diferentes estudios (Tabla 8), los valores aquí obtenidos, si bien caen dentro de éste, son en algunos casos sustancialmente diferentes a varios de tales referencias. Comparando la masa total de raíces finas de los bosques tropicales con la de los bosques templados cálidos y húmedos, Vogt, Grier y Vogt (1986), encontraron que la primera era significativamente mayor que la segunda, siendo a su vez superior la de estos últimos ecosistemas que aquella encontrada en los bosques boreales. La masa total de raíces finas aquí encontrada hasta los 80 cm de profundidad, fue de 17,77 mg ha-1 en la parcela de ciprés, 8,12 mg ha-1 en la parcela de pino pátula y 6,22 mg ha-1 en la de roble.
Tabla 8. Valores de masa total de raíces finas (0- 6 mm de diámetro) para bosques de diferentes regiones.

Los valores de ciprés y pino pátula, corresponden, en su orden, con los mencionados por Vogt, Grier y Vogt (1986) para bosques templados cálidos -19,17 mg ha-1- y para bosques templados fríos -7,31 mg ha-1- de coníferas siempreverdes, los cuales incluyen raíces finas vivas y muertas. Por otra parte, la masa de raíces finas encontrada en la parcela de roble, dista mucho de los valores encontrados por estos mismos autores para bosques latifoliados siempreverdes de los trópicos (40,68 mg ha-1) y para bosques latifoliados también siempreverdes, de la región templada (18,30 mg ha-1).
A esta situación pueden contribuir numerosos factores- sustrato edáfico, edad del rodal, etc.- sin embargo, de hecho, se ha encontrado (Vogt, Gier y Vogt, 1986) que la mayor masa de raíces encontrada en la región tropical procede no de formaciones naturales sino de plantaciones forestales establecidas con especies siempreverdes.
Cuando se compara la masa de raíces aquí obtenida con la determinada para bosques de la región templada, se encuentra que para las parcelas de roble y de pino pátula hay cierta similaridad. Al respecto, Santantonio, Hermann y Overton (1977), indican que el rango de masa de raíces finas (< 5 mm) en bosques templados de coníferas y maderas duras, varía entre 5 y 10 mg ha-1, cuando la edad del sitio supera los diez años. Mientras que los rangos para bosques templados y tropicales en general coinciden, el segundo en regiones húmedas tiende hacia el límite superior. En consonancia con lo recién mencionado, McClaugherty, Aber y Melillo (1982), registraron 9,1 mg ha-1 de raíces finas (< 3 mm de diámetro) en plantaciones de Pinus resinosa, y 10,5 mg ha-1 en bosques mixtos de maderas duras, ambos en Massachussets, EUA. Estos valores no son muy distantes de los hallados para las plantaciones de pino pátula y roble en este estudio. Al respecto, y de conformidad con los planteamientos de Baillie (1996), se ha encontrado que los bosques localizados en suelos de mayor fertilidad y con ciclos de nutrientes parcialmente abiertos, tienen profundidades de raíces similares a aquellas ubicadas en ecosistemas forestales de la región templada.
Cuando se observa la información disponible para la región tropical, se aprecia que los valores son muy disímiles entre sí. Así, para bosques de tierras bajas de la Amazonía Brasilera, la masa total de raíces finas –vivas y muertas- encontrada por Klinge (1973), osciló entre 10,9 mg ha-1 y 16,1 mg ha-1 (para 89 y 107 cm de profundidad respectivamente), mientras que para bosques secos tropicales deciduos de la India, Singh y Singh (1981) establecieron el intervalo 4,0 - 5,5 mg ha-1 hasta los 50 cm de profundidad.
Nutrientes en las raíces finas. La concentración de macroelementos –Ca, Mg, K y P- en las raíces finas fue claramente superior en las parcelas de coníferas, con respecto a las verificadas en el robledal, y siendo en la mayoría de los casos, superiores en ciprés. Para este grupo de elementos, las concentraciones halladas por Lundgren (1978) para raíces finas en plantaciones de Pinus patula y Cupressus lusitanica, fueron muy similares entre sí, siendo tales valores muy superiores a los aquí obtenidos. El hecho de que los contenidos en el suelo para este grupo de elementos en las plantaciones de Tanzania sean sustancialmente superiores a las de las parcelas en Piedras Blancas, permite sugerir que a mayor disponibilidad de nutrientes en el suelo, mayor concentración de ellos en las raíces finas. Esto guarda además correspondencia con el hecho de que una menor disponibilidad de nutrientes en el suelo, conlleva un mayor gasto de energía en la producción de raíces finas para capturar estos nutrientes y suplir las demandas nutricionales, lo cual se ve reflejado en la mayor masa de raíces finas encontradas en el área de interés en este trabajo, cuya oferta edáfica, como se ha mencionado, es claramente menor.
Los valores de concentración de nutrientes disponibles en la literatura, para las raíces finas de ecosistemas boscosos, muestran una amplia variabilidad, entre otros, aspectos tales como: el tamaño de las raíces que se consideran como finas –las cuales comprenden distintos intervalos que van desde un límite superior de 6 mm-; el tipo de clima; la clase de suelo, etc. Para un amplio número de bosques tropicales de tierras bajas (Klinge,1976; Fölster y Huber,1984; Sim y Nykvist, 1991; Bloomfield, Vogt y Vogt, 1993; Hondermann, 1995; Murach et al., 1995; Wassenaar, 1995), determinaron que las concentraciones de bioelementos vienen definidas por los siguientes intervalos: Ca (0,04-0,46 %), Mg (0,06-0,18 %), K (0,06-0,50 %) y P (0,01-0,07 %).
Siguiendo la información disponible en la literatura para la concentración de nutrientes en las raíces finas en bosques tropicales de tierras altas (Priess, Then y Fölster, 1999; Edwards y Grubb, 1982; Steinhardt, 1978), se pueden señalar los siguientes intervalos: Ca (0,11-0,73 %), Mg (0,13-0,61 %), K (0,24-0,50 %) y P (0,03-0,05 %). Como se desprende de la comparación establecida entre las concentraciones establecidas para bosques tropicales de tierras bajas y de tierras altas, los valores de estos últimos son en términos generales superiores, lo cual puede ser reflejo de unos suelos menos lixiviados y mayores niveles de fertilidad, que aquellos usualmente sometidos a regímenes de pluviosidad severos.
Al comparar las concentraciones de nutrientes presentes en las raíces de este estudio para el ecosistema natural, con las recién presentadas para el grupo de bosques montanos, se encuentra que las de estos presentan valores superiores para todos los elementos. Así, la concentración de Ca en las raíces finas del robledal -0,06 %- es extremadamente baja, reflejando la baja disponibilidad del elemento en el suelo. Dicho valor es escasamente comparable –y de lejos- con el reportado por Fölster y Huber (1984) -0,1 %- para raíces finas menores de 2 mm de diámetro, en bosques de tierras bajas de Venezuela, desarrollados sobre suelos arenosos. De manera similar al calcio, las bajas concentraciones de magnesio, las cuales alcanzan su mínimo absoluto para el conjunto de bases de cambio en las tres parcelas, sugieren la probable verificación de antagonismo entre ambos elementos.
Pasando a las plantaciones forestales, y en el caso particular de ciprés, se encuentran los máximos absolutos de calcio, y los mínimos de magnesio, situación que como antes se indicó evidencia la ocurrencia de un posible desequilibrio Ca/Mg, según el cual se ve favorecida la toma del primero de ambos elementos, ante su mayor abundancia en el compartimento edáfico y ocurrencia de valores mayores de pH en dicha parcela.
Si bien las concentraciones de potasio son claramente inferiores a las reportadas por la literatura, es necesario mencionar que el proceso de separación de las raíces del suelo, conllevó el empleo de agua a presión mediante la técnica descrita en el apartado de metodología. Así, el carácter eminentemente móvil de este elemento (Chapin, 1980) y fácil lixiviación de los tejidos vegetales, podría conllevar pérdidas sustanciales del mismo.
En cuanto a fósforo, las concentraciones extremadamente bajas encontradas en la parcela de roble, bien podrían reflejar el ampliamente reconocido proceso de inmovilización de fosfatos solubles que se da en suelos desarrollados a partir de cenizas volcánicas (Shoji, Nanzyo y Dahlgren,1993; Jackman et al., 1997) Las concentraciones de este elemento encontradas en las parcelas de coníferas se localizan cerca del límite inferior del intervalo sugerido para bosques tropicales de tierras altas, situación que podría sugerir la presencia de estructuras de tipo micorrizal en dichas plantaciones, las cuales son de crucial importancia en el ciclo del P. En la región de Piedras Blancas, Alvarado (1988 a, b), ha verificado su presencia en plantaciones de P. patula (ectomicorrizales) y de C. lusitanica (endomicorrizal).
La escasez de referencias en la literatura para el grupo de elementos menores, impide el establecimiento de comparaciones para las concentraciones en las raíces finas, siendo importante resaltar el caso de hierro y manganeso. En la parcela de roble el primero de estos alcanza el máximo absoluto de concentración, en tanto que se obtiene el mínimo absoluto de manganeso, situación que pone en evidencia la ocurrencia de antagonismo Fe/Mn. Esto corresponde, por demás, con la máxima y mínima cantidad que de Fe y Mn, respectivamente, se registran en los primeros 80 cm del suelo en el robledal.
Conforme se anotó en los resultados, los elementos Ca, Mg, K, P, Mn, Cu y Zn acumulados en las raíces finas, hasta los 80 cm de profundidad, fueron superiores en la parcela de ciprés, siendo la única excepción el microelemento Fe, el cual fue claramente superior en la parcela de roble. En las parcelas de coníferas, el patrón de abundancia siguió la secuencia decreciente Ca > K > Mg > Fe > P, siendo muy similares los valores de Fe y P en la parcela de pino pátula. Como excepción del patrón descrito, se encontró que en la parcela de roble, el elemento mayoritario fue Fe, por encima incluso de calcio, conforme se indicó para las concentraciones del elemento en esta parcela. Si bien en todas las parcelas pareciera verificarse antagonismo entre Fe y Mn, probablemente las mayores diferencias encontradas entre las concentraciones de ambos elementos en la parcela de roble, obedezcan a una influencia más marcada en las condiciones de acidez en esta última, sobre la captura preferencial de Fe sobre Mn.
Cuando se observan las cantidades totales acumuladas de elementos – Ca, Mg, K, P, Cu, Fe, Mn y Zn-, en las raíces finas, se tiene que la parcela de ciprés muestra una clara superioridad -154,55 kg ha-1- frente a la de pino pátula -38,19 kg ha-1- y la de roble -14,31 kg ha-1 – situación que no corresponde, sin embargo, con las reservas edáficas de estos elementos presentes en ellas, las cuales siguen la secuencia decreciente roble > ciprés > pino pátula, lo cual sugiere cierta independencia entre ambos compartimentos. La clara superioridad mostrada por la parcela de ciprés se explica a partir de la masa de raíces allí encontrada, la cual, es 2,2 veces mayor que la registrada en pino pátula y 2,9 veces la correspondiente a roble.
Si bien no se dispone de información experimental acerca de las tasas de producción y de descomposición de las raíces finas en este estudio, como una aproximación a la importancia potencial que éstas pueden tener en los ecosistemas estudiados, se puede acudir a la comparación de las cantidades de nutrientes acumulados en las raíces finas, y aquellas presentes en la biomasa aérea (León, 2005). Así, en la parcela de ciprés, y para el grupo de elementos aquí analizados, las cantidades presentes en las raíces finas son cerca del 11 % de aquellas presentes en la masa aérea, valor sustancialmente inferior en las parcelas de pino pátula -2,6 %- y de roble -0,7 %- con lo cual, es probable que en el retorno de bioelementos del primer ecosistema, la descomposición de raíces finas desempeñe un papel más significativo, a lo cual se sumaría, además, que en términos generales las concentraciones de elementos presentes en tales órganos vegetales, son superiores a las verificadas para roble y pino pátula.
Vogt, Gier y Vogt (1986) plantean al respecto que las concentraciones de elementos presentes en las raíces finas juegan un papel muy importante en el ciclaje de nutrientes al nivel ecosistémico, ya que las cantidades retornadas anualmente a través de ellas, podrían igualar o incluso superar las aportadas vía hojarasca foliar. Esto podría parecer contradictorio, ya que las concentraciones de nutrientes en las raíces finas son con frecuencia cercanas a la mitad de las encontradas en el follaje fresco, sin embargo, tales concentraciones permanecen estables y las tasas de retorno y subsiguiente descomposición de las raíces finas son altas (Waring y URNG, 1998).
Además, el significado que tienen las raíces finas como entradas orgánicas al suelo, ha sido ampliamente reconocido, así, se ha establecido para bosques de la región templada (Harris, Kinerson y Edwards, 1977; Persson, 1978) en que la producción de raíces pequeñas es cerca de dos veces la producción de hojarasca. Ya en términos de la descomposición y de retorno real de nutrientes al ecosistema, Vogt, Edmonds y Grier (1981); Vogt et al., (1983) han sugerido que para sitios subalpinos caracterizados por tener lentas tasas de descomposición de la hojarasca, bajos niveles de disponibilidad de nutrientes, y suelos que no se congelan durante los meses de invierno, las raíces finas pueden contribuir más que la misma hojarasca. Al respecto, y para diferentes tipos de bosques, Vogt, Grier y Vogt (1986) han reportado mayores entradas de materia orgánica al perfil edáfico a través de raíces finas que por adiciones superficiales de masa de descomposición de hojarasca, las cuales definen el intervalo 29-57 %.
A manera de síntesis, se puede afirmar que a pesar del reconocimiento cada vez mayor de la importancia que tienen las raíces finas para el crecimiento de las plantas, y en particular el papel que cumplen en los ciclos biogeoquímicos de los ecosistemas terrestres, éstas son aún pobremente conocidas. Hay quienes han considerado que las raíces contribuyen poco a los ciclos del carbono y de nutrientes con respecto a otros componentes como la hojarasca (Persson, 1990), razón por la cual en el pasado no hayan sido tenidas en cuenta. Sin embargo, se ha demostrado que las raíces finas son muy importantes para el funcionamiento del ecosistema, no sólo por su participación en la productividad primaria neta (PPN), sino también en los ciclos biogeoquímicos de los bosques, debido a que su descomposición contribuye significativamente al enriquecimiento de los horizontes orgánicos gracias a su rápida reconversión (Spurr y Barnes, 1982; Burke y Raynal, 1994). Jackson, Money y Schulze (1997) han expuesto que de la PPN en un sistema natural, las raíces finas son tal vez el más prominente sumidero de carbono, ya que se ha estimado que el reemplazo de las raíces finas por año alcanza a representar hasta un 33 % de esta productividad anual de carbono.
La importancia del estudio de las raíces finas se centra no sólo en el papel preponderante que desempeñan interviniendo directamente en la captura de bioelementos para las plantas, sino además por la alta tasa de retorno de estos al medio, aspectos ambos de crucial significado para el funcionamiento y productividad de los ecosistemas. Un primer paso lo constituye la valoración de las existencias de ellas en términos de peso y calidad en el compartimento edáfico de los ecosistemas forestales, a la cual deberán sumarse posteriormente estudios que permitan conocer aspectos propios a su dinámica tales como sus tasas de producción y de descomposición, con miras a develar capítulos funcionales hoy día muy poco conocidos en el ámbito tropical.
CONCLUSIONES Y RECOMENDACIONES 
La masa de raíces finas encontrada disminuyó muy rápidamente con la profundidad, siendo la proporción de éstas en los primeros 30 cm del suelo, máxima en la parcela de ciprés (83 %), seguida de pino pátula (75 %) y finalmente por roble (68 %). Las diferencias verificadas entre parcelas, no correspondieron con una menor disponibilidad de nutrientes que conllevara un mayor desarrollo de raíces para la exploración del suelo conducente a su captura, situación que sugiere que son otros los factores, antes que la disponibilidad de elementos nutritivos, los que controlan la proliferación de raíces finas en cercanías a la superficie.
La concentración de macroelementos en las raíces finas fue claramente superior en las parcelas de coníferas, presentándose en la mayoría de los casos, los máximos valores en ciprés. El hecho de que las concentraciones sean bajas cuando se comparan con las de otros estudios en los que se reporta una mayor oferta edáfica y una menor masa de raíces, permite sugerir que una mayor disponibilidad de nutrientes en el suelo, conlleva una mayor concentración de ellos en las raíces finas. Esta situación guarda a su vez correspondencia con el hecho de que una menor disponibilidad de nutrientes en el suelo, implica un mayor gasto de energía en la producción de raíces finas para capturar tales nutrientes, y suplir así las demandas nutricionales, lo cual se ve reflejado en la mayor masa de raíces finas encontradas en este estudio.
Las cantidades totales acumuladas de los elementos Ca, Mg, K, P, Mn, Cu y Zn en las raíces finas, fueron superiores en la parcela de ciprés, con excepción del microelemento Fe, el cual fue claramente superior en la parcela de roble. En las parcelas de coníferas, el patrón de abundancia siguió la secuencia decreciente Ca > K > Mg > Fe > P, siendo muy similares los valores de Fe y P en la parcela de pino pátula.
La secuencia descrita para las cantidades de elementos acumulados en las raíces finas, no corresponde con las reservas edáficas de los bosques estudiados, lo cual sugiere cierta independencia entre ambos compartimentos. La clara superioridad mostrada por la parcela de ciprés se explica a partir de la masa de raíces allí encontrada, la cual, es 2,2 veces mayor que la registrada en pino pátula y 2,9 veces la correspondiente a roble.
La mayoría de investigadores aceptan la importancia fisiológica que tienen las raíces finas – diámetro < 5-6 mm- en los ecosistemas forestales; sus cantidades, que varían entre 5,0 y 16,1 mg ha-1, generalmente representan cerca de la mitad del total de la biomasa de raíces pequeñas (Singh y Singh, 1981). Este hecho, sumado a aportes potenciales de elementos a través de la descomposición incluso por encima de los correspondientes a hojarasca, realza la importancia que reviste su estudio apropiado.
Es importante señalar que además de los factores físicos y biológicos que explican las diferencias en la masa de raíces disponibles en la literatura especializada, es necesario asimismo incluir la metodología empleada para el muestreo y separación. En particular pueden mencionarse, entre otros aspectos, la profundidad y la intensidad del muestreo, y la época en que éste es realizado.
Hacia el futuro, es necesario desarrollar estudios de raíces finas en ecosistemas tropicales, que sigan metodologías estandarizadas que permitan establecer comparaciones entre los resultados que se obtengan. Esta recomendación cobra especial importancia para los bosques tropicales montanos, de los cuales no se dispone de información en la materia.
Este trabajo fue realizado con el apoyo financiero de la Dirección de Investigaciones de la Universidad Nacional de Colombia, Sede Medellín y el Laboratorio de Ecología y Conservación Ambiental del Departamento de Ciencias Forestales de la misma Universidad. Los autores expresan su agradecimiento al Ingeniero Forestal Andrés Sierra Buitrago y al proyecto Dinámica del Carbono del Suelo en Bosques Tropicales Maduros y Jóvenes, por su colaboración.
ALVARADO, B. Determinación de micorrizas en Cupressus sp. En: OROZCO, F. H., ed. Investigaciones sobre micorrizas en Colombia N° 2 Medellín: Sociedad Colombiana de la Ciencia del Suelo, 1988 a. p. 111-117. [ Links ]
ALVARADO, B. Efecto de las ectomicorrizas en el desarrollo de Pinus patula. En: OROZCO, F. H., ed. Investigaciones sobre micorrizas en Colombia N° 2. Medellín: Sociedad Colombiana de la Ciencia del Suelo, 1988 b. p. 131-137. [ Links ]
BAILLIE, I. C. Soils of the humid tropics. En:. RICHARDS, P. W., ed. The tropical rain forest: an ecological study. Cambridge University Press, 1996. p. 256-286. [ Links ]
BLOOMFIELD, J. VOGT, K. A. and VOGT, D. J. Decay rate and substrate quality of fine roots and foliage of two tropical tree species in the Luquillo Experimental Forest, Puerto Rico. En: Plant and Soil. Vol. 150 (1993); p. 233-245. [ Links ]
BURKE, M. K. and RAYNAL, D. J. Fe root growth phenology, production, and turnover in a northern hardwood forest ecosystem. En: Plant and Soil. Vol. 162 (1994); p. 135-146. [ Links ]
CHAPIN, F. The mineral nutrition of wild plants. En: Annual Review of Ecology and Systematics. Vol. 11 (1980); p. 233-260. [ Links ]
CUEVAS, E. and MEDINA, E. Nutrient dynamics within amazonian forests. II. Fine root growth, nutrient availability and leaf litter descomposition. En: Oecologia. Vol. 76 (1988); p. 222-235. [ Links ]
________ and _________. Root production and organic matter decomposition in a tierra firme forest of the Upper Rio Negro Basin. En: Wurzelökologie und ihre Nutzanwendung, International Symposium Gumpenstein. Irdning, Austria , 1983. p. 653-666. [ Links ]
EDWARDS, P. J. and GRUBB, P. J. Studies of mineral cycling in a montane rain forest in New Guinea . Part 4: soil characteristics and the division of mineral elements between the vegetation and soil. En: Journal of Ecology. Vol. 70 (1982); p. 649-666. [ Links ]
EMPRESAS PÚBLICAS DE MEDELLÍN. Estudio general de coberturas vegetales de la cuenca de la quebrada Piedras Blancas. Medellín: EPM, Dirección de Planeación, Unidad de Recursos Naturales, 1989. 42 p. [ Links ]
FAHEY, T.J. et al. Fine root production and demography. En: ROBERTSON, G. P. et al., eds. Standard soil methods for long-term ecological research. Oxford, University Press, 1999. p. 437-455. (Network Series). [ Links ]
FÖLSTER, H. and HUBER, O. Interrelaciones suelos-vegetación en el área de Galipero – Territorio Federal Amazonas. Caracas: Ministerio del Ambiente y de los Recursos Naturales Renovables, 1984. 260 p. (Serie Informes Técnicos DGSIIA/IT/I44). [ Links ]
HARRIS, W. F. KINERSON, R. S. Jr. and EDWARDS, N. T. Comparison of belowground biomass of natural deciduous forests and loblolly pine plantations. En: Pedobiologia. Vol. 17 (1977); p. 369-381. [ Links ]
HERRERA, R. A. et al. Amazon ecosystems: their structure and functioning with particular emphasis on nutrients. En: Interciencia. Vol. 3 (1978); p. 223-232. [ Links ]
HOLDRIDGE, R. L. Ecología basada en zonas de vida. Costa Rica: Instituto Interamericano de Cooperación para la Agricultura, 1987. 216 p. [ Links ]
HONDERMANN, J. B. N. Fitomassa e estoque de bioelementos das diversas fases da vegetação secundária, provenientes de diferentes sistemas de uso da terra no nordeste paraense. Brasil, 1995. 184 p. Thesis (Master). Universidade Federal do Pará, Belém – PA. [ Links ]
JACKMAN, J. M. et al. Rietveld estimates of mineral percentages to predict phosphate sorption by selected Hawaiian soils. En: Soil Science Society of America Journal. Vol.61 (1997); p. 618-625. [ Links ]
JACKSON, R. B.; MOONEY, H. A. and SCHULZE, E. D. A global budget for fine root biomass, surfacea area, and nutrient contents. En: Proceedings of National Academy of Sciences. Vol. 94 (1997); p. 7362-7366. [ Links ]
KINGSBURY, N. and KELLMAN, M. Root mat depths and surface soil chemistry in southeastern Venezuela . En: Journal of Tropical Ecology. Vol. 13 (1997); p. 475-479. [ Links ]
KLINGE, H. Root biomass estimation in lowland tropical rain forests of central Amazonia, Brazil . I. Fine root masses of pale yellow latosol and a giant humus podzol. En: Tropical Ecology. Vol. 14 (1973); p. 29-38. [ Links ]
________ and HERRERA, R. Biomass studies in Amazon Caatinga forest in southern Venezuela . Part 1: standing crop of composite root mass in selected stands. En: Tropical Ecology. Vol. 19 (1978); p. 93-109. [ Links ]
KLINGE, H. IV. Nutrients in fine roots from latosols. En: Tropical Ecology. Vol. 17 (1976); p. 79-87. [ Links ]
LEÓN, J. D. Contribución al conocimiento del ciclo de nutrientes en bosques montanos naturales de Quercus humboldtii y reforestados (Pinus patula y Cupressus lusitanica) de la región de Piedras Blancas, Antioquia (Colombia). Salamanca, 2005. 98 p. Tesis (Doctoral). Universidad de Salamanca. [ Links ]
LUNDGREN, B. Soil conditions and nutrient cycling under natural and plantation forests in Tanzanian Highlands. Uppsala: Swedish University of Agricultural Sciences, 1978. 425 p. (Reports in Forest Ecology and Forest Soils; no. 31). [ Links ]
MARTYN, M. C. and ROSS, A. V. Root systems. En: PEARCY, R.W. et al., eds. Plant physiological ecology: field methods and instrumentation. Great Britain : Chapman and Hall, 1996. p. 367-371. [ Links ]
McCLAUGHERTY, C. A.; ABER, J .D. and MELILLO, J. M. The role of fine roots in the organic matter and nitrogen budgets of two forested ecosystems. En: Ecology. Vol. 63 (1982); p. 1481-1490. [ Links ]
MORENO-CHACÓN, M. and LUSK, C. H. Vertical distribution of fine root biomass of emergent Nothofagus dombeyi and its canopy associates in a Chilean temperate rainforest. En: Forest Ecology and Management. Vol. 199 (2004); p. 177-181. [ Links ]
MURACH, D. et al. Fine root inventories in Dipterocap forests and plantations in east Kalimantan, Indonesia . En: Proceedings 3rd ISSS-AISS-IBG Congress on Forest Soils, Vol. 7, Mineral nutrition Balikpapan, 1995. p. 40-53. [ Links ]
PERSSON, H. Root dynamics in young Scots pine stand in Central Sweden. En: Oikos. Vol. 30 (1978); p. 508-519. [ Links ]
PERSSON, H. Methods of studying root dynamics in relation to nutrient cycling. Methods of studying root dynamics. En: Nutrient Cycling in Terrestrial Ecosystems. 1990. p. 198-217. [ Links ]
PRIESS, J.; THEN, C. and FÖLSTER, H. Litter and fine-root production in three types of tropical premontane rain forest in SE Venezuela. En: Plant Ecology. Vol. 143 (1999); p. 171-187. [ Links ]
ROSS, S. M. Organic matter in tropical soils: current conditions, concerns and prospects for conservation. En: Progress in Physical Geography. Vol. 17 (1993); p. 265-305. [ Links ]
SANFORD, R. L. and CUEVAS, E. Root growth and rhizosphere interactions in tropical forests. En: MULKEY, S. S.; CHAZDON, R. L and SMITH, A. P., eds. Tropical forest plant ecophysiology. New York: Chapman and Hall, 1996. p. 268-300. [ Links ]
SANTANTONIO, D.; HERMANN, R. K. and OVERTON, W. S. Root biomass studies in forest ecosystems. En: Pedobiologia. Vol. 17 (1977); p. 1-31. [ Links ]
SHOJI, S.; NANZYO, M. and DAHLGREN, R. A. Volcanic ash soils-genesis, properties, and utilization. Amsterdam: Elsevier, 1993. 312 p. [ Links ]
SIERRA, A. Producción y descomposición de raíces finas en bosques tropicales maduros y jóvenes de la región de Porce. Medellín, 2004. 43 p. Trabajo de grado (Ingeniero Forestal). Universidad Nacional de Colombia. Facultad de Ciencias Agropecuarias. [ Links ]
SIM, B. L. and NYKVIST, N. Impact of forest harvesting and replanting. En: Journal of Tropical Forest Science. Vol. 3 (1991); p. 251-284. [ Links ]
SINGH, K. P. and SINGH, R. P. Seasonal variation in biomass and energy of small roots in tropical dry deciduous forest, Varanasi, India . En: Oikos. Vol. 37 (1981); p. 88-92. [ Links ]
SPURR, S. H. y BARNES, B. V. Ecología forestal. México: A.G.T., 1982. 538 p. [ Links ]
St. JOHN, T. V. Response of tree roots to decomposing organic matter in two lowland Amazonian rain forests. En: Canadian Journal of Forest Research. Vol. 13 (1983); p. 346-349. [ Links ]
STARK, N. M. and JORDAN, C. F. Nutrient retention by the root mat of an Amazonian rain forest. En: Ecology. Vol. 59 (1978); p. 434-437. [ Links ]
STARK, N. and SPRATT, M. Root biomass and nutrient storage in rain forest oxisols near San Carlos de Rio Negro. En: Tropical Ecology. Vol. 18 (1977); p. 1-9. [ Links ]
STEINHARDT, U. Untersuchungen über den Wasser- und Nährstoffhaushalt eines andinen Wolkenwaldes in Venezuela . Göttingen, 1978. 185 p. Thesis (Ph.D.). University of Göttingen. [ Links ]
USME, P. Demografía de raíces finas en bosques primarios y secundarios tropicales en la región de Porce. Medellín, 2004. 64 p. Trabajo de grado (Ingeniero Forestal). Universidad Nacional de Colombia. Facultad de Ciencias Agropecuarias [ Links ]
VOGT, K. A. EDMONDS, R. L. and GRIER, C. C. Seasonal changes in biomass and vertical distribution of micorrhizal and fibrous-textured conifer fine root in 23- and 180- year-old subalpine Abies amabilis stands. En: Canadian Journal of Forest Research. Vol. 11 (1981); p. 223-229. [ Links ]
________ et al. Organic matter and nutrient dynamics in forest floors of young and mature Abies amabilis stands in western Washington, as affected by fine-root input. En: Ecological Monographs. Vol. 53 (1983); p. 139157. [ Links ]
________; GRIER, C. G. and VOGT, D. J. Production, turnover and nutrient dynamics of above- and belowground detritus of world forests. En: Advances in Ecological Research. Vol. 15 (1986); p. 303-377. [ Links ]
________. et al. Roots and mycorrhizas in plantation ecosystems. En: CAMBIAR, E. K. S. and BROWN, A.G., eds. Management of soil, nutrients and water in tropical plantation forests. Camberra, Australia : ACIAR/CSIRO/CIFOR. ACIAR, 1997. p. 247-296. [ Links ]
WARING, R. H. and RUNNING, S. W. Forest ecosystems. Analysis at multiple sacales. San Diego, California: Academic Press, 1998. 370 p. [ Links ]
WASSENAAR, T. Roots, their dynamics and distribution. A study on standing stocks and fine root dynamics in Colombian Amazonia. Bogotá, Colombia: The Tropenbos Foundation, 1995. 54 p. [ Links ]

