










Services on Demand
Journal
Article
 Spanish (pdf)
Spanish (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mailIndicators
-
 Cited by SciELO
Cited by SciELO -
 Access statistics
Access statistics
Related links
-
 Cited by Google
Cited by Google -
 Similars in
SciELO
Similars in
SciELO -
 Similars in Google
Similars in Google
Share

 Permalink
PermalinkRevista Facultad Nacional de Agronomía Medellín
Print version ISSN 0304-2847
Rev. Fac. Nac. Agron. Medellín vol.60 no.1 Medellín Jan./June 2007
CICLAJE Y PÉRDIDA DE NUTRIENTES DEL SUELO EN BOSQUES ALTOANDINOS DE ANTIOQUIA, COLOMBIA
NUTRIENT CYCLING AND NUTRIENT LOSSES IN ANDEAN MONTANE FORESTS FROM ANTIOQUIA, COLOMBIA
Adriana Londoño Álvarez1; Diana Cristina Montoya Gómez2; Juan Diego León Peláez3 y Maria Isabel González Hernández4
1 Ingeniera Forestal. Universidad Nacional de Colombia, Sede Medellín. Facultad de Ciencias Agropecuarias. A.A. 1779, Medellín, Colombia. <ala2x@yahoo.es>
2 Ingeniera Forestal. Universidad Nacional de Colombia, Sede Medellín Universidad Nacional de Colombia Sede Medellín. Facultad de Ciencias Agropecuarias. A.A. 1779, Medellín, Colombia. <dianamg2x@yahoo.es>
3 Profesor Asociado. Universidad Nacional de Colombia, Sede Medellín. Facultad de Ciencias Agropecuarias. A.A. 1779, Medellín, Colombia. <jdleon@unalmed.edu.co>
4 Profesora Titular. Universidad de Salamanca, Facultad de Ciencias Agrarias y Ambientales. Avd. Filiberto Villalobos, 119.37007, Salamanca, España. <mimg@gugu.usal.es>
Recibido: Julio 7 de 2006; aceptado: marzo 16 de 2007.
RESUMEN
El agua gravitacional y su composición química fueron medidos en bosques montanos de Quercus humboldtii y reforestados (Pinus patula y Cupressus lusitanica) de la región de Piedras Blancas, Antioquia (Colombia), por un período de tiempo de dos años. Se utilizaron lisímetros sin tensión con el fin de estimar el agua gravitacional y los flujos de nutrientes a diferentes profundidades en el perfil del suelo. El mayor valor anual de agua gravitacional en el nivel más profundo (50- 80 cm), fue hallado en la cobertura de ciprés ( 492,7 mm), seguido por pino pátula ( 14,2 mm) y roble ( 2,0 mm). De manera similar ocurrió con la pérdida de nutrientes, mostrando el mismo patrón hallado para el agua gravitacional. Así, para roble, pátula y ciprés, en su orden, se presentaron los siguientes valores de pérdida: Ca: 0,004, 0,084 y 2,270 kg ha-1 año-1; P: 0,008, 0,052 y 1,234 kg ha-1 año-1; Mg: 0,004, 0,022 y 0,667 kg ha-1 año-1. De K se registraron 0,08 y 7,092 kg ha-1 año-1 para roble y ciprés respectivamente. Estos flujos siguieron el siguiente orden según cobertura, roble: K>P>Ca>Mg, pátula: Ca>Fe>P>Mg>Zn>Mn, y ciprés: K>Mn>Ca>P>Fe>Zn>Mg.
Palabras claves: Cupressus lusitanica, Pinus patula, Quercus humboldtii, bosques montanos tropicales, plantaciones forestales tropicales, ciclo de nutrientes.
ABSTRACT
Gravitational flow and its chemical composition were measured in montane oak forests (Quercus humboldtii), in pine (Pinus patula) and cypress (Cupressus lusitanica) plantations in Piedras Blancas, Antioquia ( Colombia), over two years. Zero tension lysimeters were used at different depth soil levels. The highest gravitational flow value at highest depth (50- 80 cm) was obtained in cypress plot ( 492,7 mm), followed by pine ( 14,2 mm) and oak forest ( 2,0 mm). A similar behavior was encountered for nutrient losses, following the same pattern as gravitational flow. Thus, for oak, pine and cypress, nutrient losses were respectively: Ca: 0,004, 0,084 and 2,270 kg ha-1 y-1; P: 0,008, 0,052 and 1,234 kg ha-1 y -1; Mg: 0,004, 0,022 and 0,667 kg ha-1 y-1. K losses were 0,08 and 7,092 kg ha-1 y-1 for oak forest and cypress plantation respectively. Nutrient losses followed the next order for each type of forest: oak: K>P>Ca>Mg, pine: Ca>Fe>P>Mg>Zn>Mn, and cypress: K>Mn>Ca>P>Fe>Zn>Mg.
Key words: Cupressus lusitanica, Pinus patula, Quercus humboldtii, tropical montane forests, tropical forest plantations, nutrient cycling.
 MÉTODOS
MÉTODOS
RESULTADOS Y DISCUSIÓN
CONCLUSIONES
RECONOCIMIENTOS
BIBLIOGRAFÍA
En los ecosistemas terrestres el compartimento edáfico constituye el principal medio a través del cual se viabiliza el retorno de nutrientes a la vegetación en pie. El ciclaje y pérdida de nutrientes del suelo como procesos en ecosistemas forestales, han demostrado ser críticos para su adecuado funcionamiento. Así, pérdida de nutrientes del sistema, interrupción de los procesos de ciclaje y captura de nutrientes, o desequilibrios en el estado nutricional podrían estar asociados con declinaciones en la productividad y estabilidad forestal (Nilsson et al. 1995).
En los suelos forestales es frecuente que gran parte del agua fluya a través de las vías de circulación rápida, las cuales se corresponden con los macroporos del suelo originados principalmente por raíces muertas y por actividad animal, con lo cual se minimizan las vías de circulación capilar (Voigt 1980, Beven y German 1982, Moreno et al. 1996).
Las concentraciones de elementos presentes en la precipitación incidente que constituyen los valores referenciales para evaluar las modificaciones del agua a su paso por el bosque, varían entre coberturas a medida que dicho flujo atraviesa el dosel, cae al piso forestal y se infiltra en el suelo. Las concentraciones de la solución que alcanzan el suelo dependen, fundamentalmente, de la calidad de la precipitación y de los procesos que ocurren durante su transferencia al dosel (deposición oculta y seca y lixiviación) (Ranger, Marques y Jussy 2001).
La composición de la solución edáfica, es el resultado de procesos complejos tales como las entradas de los elementos de las soluciones gravitacionales (la lluvia y su posterior redistribución), la producción interna de elementos por mineralización de la materia orgánica del suelo y la meteorización, complejos equilibrios de iones intercambiables y las salidas por absorción (raíces y microbiota) y por drenaje profundo. La intensidad de cada proceso depende de muchos factores (profundidad del suelo, movilidad de las soluciones, requerimientos del rodal, entre otros) y los mecanismos individuales dificultan la identificación de interacciones entre ellos (Zabowski y Ugolini 1990, Marques y Ranger 1997, Ranger, Marques y Jussy 2001).
El objetivo de esta investigación comprende la evaluación y comparación del ciclaje y pérdida de nutrientes solubles en las aguas gravitacionales a diferentes profundidades del suelo, en ecosistemas forestales de roble (Quercus humboldtii Bonpl.), pino pátula (Pinus patula Schltdl. & Cham.) y ciprés (Cupressus lusitanica Mill.) de la región de Piedras Blancas (Antioquia, Colombia), por un período de dos años.

Área de estudio. La zona de estudio se localiza al oriente del departamento de Antioquia (Colombia), cuenca hidrográfica de la quebrada Piedras Blancas, siendo su altitud aproximada de unos 2.490 m. Geográficamente está ubicada en las coordenadas planas del sistema nacional cartográfico (Plancha IGAC 147-I-D) Y: 840.500 – 841.500, X: 1.188.000 – 1.189.000. La temperatura media anual es de 14,9 ºC, la humedad relativa promedio anual 82 %, y la precipitación media anual, período 1942-1997, de 1.698 mm1. Las condiciones climáticas señaladas corresponden a
la zona de vida bosque húmedo Montano Bajo (bh-MB) (Holdridge 1987). La fisiografía se caracteriza por colinas de pendiente media y suelos derivados de ceniza volcánica.
Métodos de campo y laboratorio. Entre los meses de febrero y marzo de 2001 se establecieron tres (3) parcelas permanentes de 0,5 ha en bosques de roble, pino pátula y ciprés, dotadas con dispositivos de medición para el estudio y registro de variables climáticas y procesos del ciclo biogeoquímico. En particular se colocaron pluviómetros por fuera y dentro del bosque de seis pulgadas de diámetro dispuestos 1m sobre la superficie del terreno, con miras a conocer la cantidad y la composición química de la precipitación interna (PI) y del agua proveniente por precipitación incidente. La distancia entre parcelas no superó los 200 m una de otra, siendo en esencia iguales las características del medio físico y climático. Algunas características de estos bosques se presentan en la Tabla 1.
Tabla 1. Caracterización de los rodales en la cuenca hidrográfica de la quebrada Piedras Blancas (Antioquia, Colombia).

Para el estudio de la composición química de las aguas gravitacionales a diferentes profundidades del suelo, en cada una de las parcelas referidas se abrieron dos calicatas (2x1 m) a media ladera. Éstas se dispusieron en un relieve quebrado con pendientes fuertes -50 %- sobre un manto de cenizas volcánicas con anfibolitas como material subyacente. El régimen de humedad del suelo es údico con un desarrollo efectivo de profundo a muy profundo y un drenaje natural bueno. Los suelos pertenecen a los grandes grupos Fulvudands y Hapludands de acuerdo con el sistema taxonómico de la USDA (Soil Survey Staff 2003) y Andosol úmbrico según la World Reference Base for Soil Resources (FAO 2006).
Las características edáficas son bastante homogéneas en las tres parcelas, con ligeras diferencias en la de ciprés respecto a las de roble y pino pátula pero que no afectan a su clasificación. La Tabla 2 muestra las propiedades químicas de los perfiles del suelo representativos de cada cobertura.
Tabla 2. Propiedades químicas de los suelos en las parcelas estudiadas en la cuenca hidrográfica de la quebrada Piedras Blancas (Antioquia, Colombia).

Para la valoración del ciclaje de nutrientes en la solución edáfica se utilizó la técnica de lisimetría sin tensión. La necesidad de definir el tipo de agua recolectada es un aspecto que determina la elección de la clase de lisímetro a utilizar en un estudio. Los lisímetros con tensión recogen agua retenida a potenciales difíciles de conocer y con una pauta un tanto aleatoria, mientras que los lisímetros sin tensión sólo recogen agua que percola a través del perfil, es decir, agua insaturada (Haines, Waide y Todd 1982), siendo esta última, la que constituye la principal forma de pérdida de agua del suelo.
El agua en el suelo se encuentra adherida a partículas sólidas en forma de película y de solución. Dicha adhesión conduce a estimar la retención de humedad en términos de fuerzas que son ejercidas sobre el agua en el suelo, conocidas como Tensión de Humedad o Potencial Total del Agua, que a su vez están conformadas por diversos componentes del suelo, como sólidos (potencial matricial), solutos (potencial osmótico), gravedad (potencial gravitacional) y presión (potencial de presión) (Gavande 1972, Jaramillo 2002).
El potencial más importante en las relaciones suelo-agua-planta consta de un componente matricial (m) y un componente osmótico (p):

donde el subíndice s significa esfuerzo (succión), término que se emplea comúnmente para denotar el potencial asociado con el aprovechamiento del agua por las plantas (Baver, Gardner y Gardner 1973). El agua no solamente se infiltra y almacena para uso futuro, sino también se mueve de un suelo no saturado a las raíces de la planta, y a los sitios de evaporación, o se pierde por penetración profunda. El flujo de agua en un suelo insaturado puede ocurrir también como una película de arrastre a lo largo de las paredes de los poros anchos ó como un flujo tubular a través de los poros estrechos llenos de agua. Los dos tipos de flujo pueden, de hecho, ocurrir conjunta y simultáneamente en dominios adyacentes dentro del suelo (Hillel 1998).
Quizá la diferencia más importante entre los flujos saturado e insaturado del agua en el suelo radica en su conductividad hidráulica. Cuando el suelo está saturado los poros están llenos de agua y conduciendo, siendo la fase hídrica continua y la conductividad máxima. Al suelo desaturarse algunos de los poros se llenan de aire de forma que la porción conductiva del suelo disminuye; con-forme se desarrolla la succión, los primeros poros en vaciarse son los de mayor tamaño, los cuales son los más conductivos, relegándose así a los poros de menor tamaño (Hillel 1998).
Con el fin de monitorear las aguas gravitacionales del suelo en cada una de las calicatas mencionadas se instalaron ocho lisímetros plásticos sin tensión, a razón de dos según criterio de profundidad. Estos dispositivos cumplen los principios de un lisímetro ideal (Keller y Vedy 1991, Moreno 1994), consistente en un dispositivo sin tensión, abierto, de pequeña talla e inerte desde el punto de vista físico-químico. Físicamente consisten en bandejas de 20 x 30 cm seccionadas e introducidas en el suelo siguiendo la dirección de la pendiente topográfica, conectadas a tarros de plástico de alta densidad, mediante mangueras plásticas, a través de los cuales es posible recoger tanto el flujo vertical como el horizontal, y tienen una escasa demanda de mantenimiento. Estos dispositivos fueron distribuidos en el perfil del suelo de la siguiente forma: N0, correspondiente a la primer capa del suelo que comprende el material orgánico en descomposición, N1 ubicado en un rango de profundidad de 0 a 30 cm, N2 de 30 a 50 cm y N3 a una profundidad mayor a 50 cm.
Los muestreos volumétricos se realizaron semanalmente, y mensualmente practicada una determinación química, sobre una muestra combinada de cada nivel por cobertura. El número total de semanas muestreadas, sobre las que se practicaron análisis químicos, fue de 22 para los dos años de estudio. Para la determinación de calcio (Ca), magnesio (Mg) y potasio (K) se procedió a hacer lectura directa sobre el agua previamente filtrada en papel banda azul, usando espectrofotometría de absorción atómica (Perkin Elmer 2380), previa preparación de estándares.

Figura 1. Batería de lisímetros a cero tensión a diferentes profundidades en el perfil del suelo. (Adaptada de Moreno 1994).
El análisis de hierro (Fe), manganeso (Mn) y zinc (Zn) requirió la concentración del agua, de manera que 200 ml fueron llevados a 25 ml. A la solución concentrada se le adicionó H2SO4 y HNO3, calentando a 100 ºC hasta la aparición de humo blanco. La solución resultante se filtró con papel banda azul y se llevó a 25 ml usando agua destilada para la realización de la lectura mediante espectrofotometría de absorción atómica.
La determinación de fósforo (P) se realizó siguiendo el método del ácido ascórbico. Para el efecto se tomaron 50 ml de agua para ser sometida a digestión con H2SO4 y HNO3 durante aproximadamente 30 minutos. Posteriormente se llevó la muestra a 50 ml neutralizándola con NaOH. Usando fenoltaleína como indicador, la muestra se llevó al espectro-fotómetro ultravioleta visible (Bausch and Lomb Milton Roy 601) para una lectura a 660 nm, usando el blanco correspondiente como solución de referencia. No fue necesaria la corrección por turbidez, dado que las aguas recolectadas no presentaron este fenómeno.
Los cálculos anuales comprendieron, en el caso de la concentración de bioelementos, la consideración de las cantidades de agua recogida en el flujo, con el fin de obtener la concentración media ponderada de cada elemento por evento de muestreo, por año. A partir del producto de este dato con la magnitud del flujo en cada año por nivel, fueron obtenidas las cantidades totales en ciclaje y pérdida de cada elemento.
Flujo de nutrientes. En la Tabla 3 se muestran los valores medios anuales de las aguas gravitacionales por cobertura y nivel. Los detalles pormenorizados del movimiento gravitacional de las aguas en el suelo y sus relaciones con las variables climáticas y físicas en el período de estudio no se incluyen en este trabajo ya que han sido previamente publicados (Londoño et al. 2005), por lo cual se retoman de ella únicamente los datos necesarios para el cálculo de los flujos de nutrientes. Así, a partir de los valores medios del agua gravitacional (AG), y de las concentraciones medias ponderadas por volumen de cada elemento, han sido calculados los flujos medios anuales de macro y micro nutrientes.
Tabla 3. Agua gravitacional promedio anual según cobertura y nivel (mm) en bosques altoandinos de Antioquia, Colombia.

Dado que los nutrientes que alcanzan el piso del bosque a través del flujo de Precipitación Interna (PI: agua lluvia que atraviesa el dosel habiendo o no entrado en contacto con éste, y que se computa como ingreso hasta el piso forestal dado que es recogida por pluviómetros ubicados dentro del bosque), constituyen los valores referenciales para observar su evolución conforme el agua atraviesa el mantillo y posteriormente los horizontes minerales del suelo, estos se presentan en la Tabla 4. Por su parte, las Tablas 5 y 6 contienen los flujos medios anuales de macro y micro nutrientes respectivamente.
Tabla 4. Flujo medio de los macro y micro nutrientes a través de la Precipitación Interna (PI) en bosques altoandinos de Antioquia, Colombia.
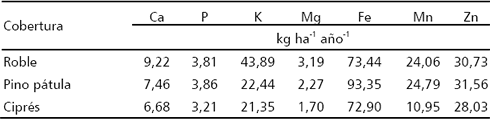
Tabla 5. Flujo medio anual de macro nutrientes (kg ha-1 año-1) en el agua gravitacional según cobertura y nivel, en bosques altoandinos de Antioquia, Colombia.

Tabla 6. Flujo medio anual de micro nutrientes (kg ha-1 año-1) en el agua gravitacional según cobertura y nivel, n bosques altoandinos de Antioquia, Colombia.

Los flujos de nutrientes a través del AG se encuentran claramente influenciados por los correspondientes a los de la PI, ya que éstos representan un vector fundamental de nutrientes al sistema hasta su llegada al suelo forestal. A este nivel, se presenta una interacción de gran relevancia entre dichas aguas y los diferentes horizontes del suelo, de la cual se derivará nuevamente, o bien una ganancia o una pérdida de nutrientes a través de las diferentes profundidades en el perfil edáfico.
Para la totalidad de macro elementos, se observa claramente una disminución sensible de ellos en el AG con respecto al agua de PI, la cual puede deberse parcialmente a la pérdida potencial de nutrientes vía escorrentía superficial. Así, a pesar de que existe un enriquecimiento por los procesos de descomposición y mineralización de la materia orgánica en el mantillo, y se encuentra disponible una gran cantidad de formas solubles de los elementos, éstas pueden ser lavadas por el flujo de escorrentía superficial impidiendo su llegada hasta la solución del suelo. En los sitios de estudio de hecho fueron verificados valores de escorrentía superficial inusualmente altos, caso de la cobertura de ciprés para la cual Ruiz, Acosta y León 2005, reportaron hasta 11,05 % del total del agua lluvia convertida en dicho flujo, el cual no sólo era determinado por el proceso de repelencia al agua desarrollado en el horizonte A del suelo (verificado por Arbeláez et al. 2004), sino que éste además le imprimía un modelo alterado para la circulación interna del agua en el suelo, conforme se desprende del patrón irregular de las aguas gravitacionales con el incremento en profundidad, expresado en la Tabla 3, el cual ya ha sido discutido detalladamente por Londoño et al. 2005.
De otro lado, ha sido reportada por distintos autores (Herrera et al. 1978, Cuevas y Medina 1988) como una posible fuente de pérdida, aquella ocurrida por la toma directa de algunos nutrientes - K, Ca, Mg y P- por las raíces, desde el material acumulado sobre el piso y sometido al proceso de descomposición. Sin embargo, al intentar explicar tales disminuciones en la cantidad de nutrientes vía toma directa por las raíces del mantillo, no se encuentra una clara asociación entre la masa total de raíces finas presente en la hojarasca acumulada sobre el piso forestal en cada cobertura, y las disminuciones encontradas experimentalmente. Así, calculando el índice de retención del mantillo (Irm) como la diferencia en las cantidades presentes de cada elemento en PI y AG, con respecto a las presentes en PI, no fueron encontradas diferencias estadísticamente significativas (P>0,01) entre las coberturas para ninguno de los elementos. A pesar de que en el mantillo de roble se registra el mayor peso seco de raíces finas (3,9 Mg ha-1), es en éste donde los valores del Irm son más bajos, encontrándose el mínimo absoluto allí para Mg (0,73 o 73 %).
Para las aguas que atraviesan el mantillo (N0), roble mostró el mayor valor en el flujo de nutrientes, evidenciándose así una relación de proporcionalidad directa entre los volúmenes de agua y los flujos de bioelementos, ya que en esta parcela es donde se tienen los máximos valores de agua colectada para N0 ( 327,82 mm). Es probable que los valores de retención del mantillo recién discutidos sean menores en este ecosistema como resultado de una capa de hojarasca que no representa un obstáculo fuerte para impedir el paso del agua y nutrientes en solución a través suyo, y consecuente llegada a los horizontes minerales.
En esta misma cobertura se observa además (Tabla 4), cómo los valores de macro nutrientes en PI son mayores en comparación con los de este flujo en las plantaciones forestales de coníferas, lo que se traduce en una mayor fuente de enriquecimiento para las aguas recolectadas en N0. Consecuentemente, las aguas que atraviesan el mantillo tienen en todos los casos mayores cantidades de bioelementos en el robledal que en las plantaciones de pino pátula y de ciprés.
Pasando ya del horizonte 0 al suelo mineral, cuando se observan los flujos de macro nutrientes (Tabla 5), se encuentra que en las tres coberturas estos muestran relación con las cantidades de AG en cada una de las profundidades monitoreadas. Así, en el robledal, los mayores flujos de bioelementos se presentaron en aquellos niveles que generaron los mayores valores de AG, siguiendo todos ellos un patrón de proporcionalidad directa con los volúmenes colectados. De manera análoga ocurrió en la cobertura de pino pátula para el total de los casos -macro y micro nutrientes- conservándose la proporcionalidad descrita en las diferentes profundidades. En la cobertura de ciprés se presentó al igual que en las anteriores un patrón de proporcionalidad directa entre el flujo de macro elementos y el AG, situación característica de una fuente sólida muy soluble la cual satura la solución (Arias et al. 2002).
Al comparar las coberturas entre sí, se tiene que ciprés presenta un comportamiento singular, ya que en ella, N3 es quien mayor cantidad de AG presenta, seguido por N2, N1 y finalmente por N0 (Tabla 3); en contraste con roble y pino pátula, que siguen un patrón general de recolección inverso al presentado por ciprés, y que corresponde al usualmente encontrado de disminución de la cantidad de agua gravitacional a medida que la profundidad aumenta; en ciprés, además, se presentaron los máximos absolutos de AG. En las tres coberturas se encontró la relación de proporcionalidad directa entre AG y flujos de bioelementos en los diferentes niveles de profundidad del suelo mineral. Así, los mayores flujos de bioelementos se presentaron en la cobertura de ciprés, seguida por la de pino pátula, en donde los valores de AG fueron intermedios; y finalmente por la de roble, en la cual los valores de AG fueron los más bajos.
En los suelos tropicales fuertemente meteorizados y escasos de nutrientes, las concentraciones de elementos en el lixiviado del mantillo son generalmente más altas que en la solución del suelo mineral, debido a una mayor concentración de nutrientes en el horizonte orgánico que en los horizontes minerales (Bruijnzeel et al. 1993, Lilienfein et al. 2000). En los suelos minerales los nutrientes son absorbidos por las plantas, dando como resultado menores concentraciones en la solución, de donde pueden derivarse los valores hallados para los niveles 1, 2 y 3 para las coberturas de roble y pátula donde del mantillo del piso del bosque se derivan mayores flujos de elementos por la hojarasca característica de estas coberturas.
Por otra parte, el hecho de que las concentraciones de los elementos en la solución del suelo sean menores que en el lixiviado del mantillo, indica que posiblemente la mayoría de los elementos es tomada por la vegetación o retenida por el suelo (Zech, Wilcke y Valarezo 1999). Adicionalmente, el componente principal del balance general es el ciclo absorción/acumulación de hojarasca, de manera que la capa superficial presenta una tasa de enriquecimiento lo que supone una pérdida de los mismos en las capas más profundas para el caso de los elementos poco móviles (Gómez-Rey y De Anta 2002).
Se sabe que las propiedades químicas edáficas, influyen en las concentraciones de los componentes de la solución del suelo; en cuanto al pH (Kamprath 1980) muchos de los suelos de los trópicos húmedos son ácidos e incluso fuertemente ácidos. Para este estudio, los suelos se hallan en un ámbito ácido con valores de pH de 4,0 a 5,0. Comúnmente la acidez del suelo indica la existencia de niveles bajos de los cationes principales: Ca, Mg y K (Fassbender, Hans y Bornemisza 1987), lo que está relacionado a su vez con los valores relativamente bajos encontrados en la solución del suelo monitoreada. Esta solución fue así mismo ácida, y aún cuando es impreciso definir un patrón, podría señalarse una tendencia leve hacia el aumento de pH a medida que atraviesa los horizontes minerales. De igual forma coincide la acidez del suelo con la de la solución, obteniéndose los mayores valores de pH en la solución de la cobertura de ciprés (Tabla 7).
Tabla 7. Comportamiento del pH en el agua gravitacional según cobertura y nivel, en bosques altoandinos de Antioquia, Colombia.

En los flujos estudiados se registró el patrón usualmente reportado de dominancia de Ca sobre Mg, si bien la relación Ca/Mg fue relativamente baja, oscilando entre 1,5 y 3,4 tomando como base los valores medios. La movilidad catiónica en este estudio aumentó en el orden Mg<Ca<<K, presentándose en la generalidad de los casos los mayores valores de movilidad de Ca en la parcela de ciprés. De acuerdo con Jaramillo 1995, tal parece que en los Andisoles del Oriente Antioqueño, existe selectividad en la retención de Mg por el suelo lo que le hace menos susceptible a la lixiviación que los demás cationes básicos.
El elemento de mayor participación dentro de los cationes de la solución del suelo en las tres coberturas fue el potasio, a diferencia del patrón comúnmente exhibido en este aspecto por el calcio, el cual se presentó como catión principal en la mayoría de los suelos, situación atribuible al importante efecto de lavado que la lluvia hace de K de los tejidos foliares, y que en forma de PI enriquecida por este elemento, alcanza el piso forestal.
En cuanto a P, los suelos derivados de cenizas volcánicas poseen una gran capacidad de fijación de fosfatos solubles, que se relacionan con la poca disponibilidad del elemento en los suelos del Oriente Antioqueño (Tschinkel 1972). Dado el pretratamiento de digestión ácida aquí practicado sobre las muestras de agua, los valores de P indicados comprenden además de los fosfatos solubles, el P presente en el material particulado en solución –estructuras vegetales, hongos, bacterias, partículas de arcilla- situación que conlleva incrementos en los valores de flujo estimados para este estudio. Así, estos flujos se muestran claramente superiores a aquellos reportados en la literatura disponible.
En términos de pérdidas de bioelementos solubles para el sistema, es decir, aquellos que como drenaje profundo no alcanzan a ser tomados por las raíces de las plantas, fue necesario establecer un nivel de profundidad que definiera tal situación. Dado que la concentración de raíces finas se hace mínima en las tres coberturas a la profundidad de 50- 80 cm, ha sido aquí propuesto N3 como nivel de referencia para computar las pérdidas. Al respecto, para las tres parcelas en estudio, Barreto y León 2005, reportaron un rápido descenso en la masa de raíces finas con el incremento en profundidad, encontrándose en la parcela de ciprés el 83 % de las raíces en los primeros 30 cm de profundidad, en pino pátula el 75 % y en roble el 68 %. Para el nivel de referencia mencionado (N3) la cantidad de raíces finas alcanza su máximo absoluto en la cobertura de pino pátula con 8,7 %, seguido de roble con 6,6 % y finalmente de ciprés con 2,7 %.
Para el conjunto de macro elementos las mayores cantidades computadas como pérdidas para el sistema se presentan en las parcelas de coníferas, en donde las láminas de agua gravitacional son claramente mayores que las encontradas en la parcela de roble, situación que incide directamente en el cálculo del flujo a esta profundidad y que se hace especialmente notoria en la parcela de ciprés. Al respecto, ha sido ampliamente expuesto que en un bosque maduro los nutrimentos son mantenidos en un ciclo casi cerrado, del cual pocos nutrientes se pierden en el drenaje. Así, las mayores pérdidas de Ca y Mg por drenaje profundo se encontraron en ciprés, a las cuales siguieron en su orden pino pátula y finalmente roble. Los valores de pérdida aquí obtenidos son inferiores a los reportados para estos elementos en otros bosques tropicales, conforme puede apreciarse en la Tabla 8.
Tabla 8. Valores reportados de pérdida de macro nutrientes para la región tropical por diferentes métodos.

Del grupo de elementos estudiados fue K el que mostró las máximas pérdidas, siendo a su vez la cobertura de ciprés en donde se encontró el máximo absoluto. Los altos valores de pérdida de este elemento pueden deberse a su carácter eminentemente móvil, encontrándose en las plantas en forma K+ -sin formar ningún compuesto- y propiciándose su lavado de los tejidos foliares por la precipitación, llegando al piso del bosque mediante las aguas de escurrimiento cortical y precipitación interna, de donde es aparentemente lavado con facilidad por la incapacidad del complejo de cambio para retenerlo. En comparación con otros estudios (Tabla 8), la pérdida de K de esta cobertura se encuentra dentro de los valores reportados para bosques húmedos tropicales.
La pérdida de P en pino pátula 0,05 kg ha-1 año-1, se corresponde de cerca con la encontrada por Russell 1983, 0,04 kg ha-1 año-1, en bosques tropicales de tierras bajas de Brasil, en tanto que la pérdida de este elemento en ciprés 1,23 kg ha-1 año-1, es cercana a la mitad de la encontrada por Huttel 1975, 2,2 kg ha-1 año-1, en bosques tropicales también de tierras bajas, de Costa de Marfil. En el caso de los micronutrientes, no se encontraron valores reportados para bosques tropicales que permitieran comparar las pérdidas observadas en este estudio. Los oligoelementos aquí estudiados (Fe, Mn y Zn), corresponden con los de carácter catiónico, cuya solubilidad se encuentra fuertemente influenciada por el pH. Así, de estos tres micro elementos, fue Fe quien presentó las mayores concentraciones en el AG, mostrando Mn y Zn valores semejantes. Tales valores de Fe se relacionan con los de pH bajos encontrados en la solución.
Según Vitousek y Sanford 1986, las pérdidas de nutrientes son probablemente mayores en zonas más fértiles; sin embargo, antes de definir un patrón es necesario obtener información de experimentos desarrollados por períodos de tiempo representativos. De este estudio se desprende que al parecer, el ecosistema forestal natural hace un uso más eficiente de los nutrientes en solución, que el correspondiente a las plantaciones forestales de coníferas; en gran medida esta situación está ligada a una mayor eficiencia en el uso del agua por el robledal, ya que de la observación de las características químicas del suelo no se puede concluir acerca de la verificación de un proceso de acumulación de bioelementos en él.
Para el conjunto de elementos estudiados -en las diferentes coberturas y niveles- se encontró un efecto neto de disminución de ellos con respecto a las cantidades presentes en la precipitación interna, situación promovida parcialmente por la verificación de procesos de retención en el mantillo y por acción de la escorrentía superficial, impidiendo su llegada efectiva al suelo.
Las cantidades de nutrientes asociadas a las aguas gravitacionales, estuvieron claramente relacionadas con las cantidades de agua gravitacional. Así, siguieron la secuencia decreciente ciprés>pino pátula> roble, lo cual se verificó para los diferentes niveles entre coberturas.
Las mayores pérdidas de bioelementos se encontraron en la plantación de ciprés, situación claramente asociada a la ocurrencia de los máximos absolutos de las aguas gravitacionales en el tercer nivel de profundidad en dicho ecosistema.
El patrón de pérdida de nutrientes encontrado, permite señalar de mayor a menor, los ecosistemas de ciprés, pino pátula y roble, lo cual sugiere una economía de nutrientes más eficiente en el caso de la cobertura natural, con respecto a aquella registrada en las plantaciones forestales. Los valores de pérdida de macro elementos encontrados, corresponden con los límites inferiores de los rangos reportados por otros estudios realizados en la región tropical, lo cual puede deberse a los bajos contenidos y/o disponibilidad de estos en el suelo.
Los estudios realizados mediante empleo de lisímetros, permiten un mejor conocimiento de los procesos que ocurren en el sistema edáfico, especialmente en la zona radical; sin embargo, diversos autores precisan que los volúmenes de los flujos de agua sean estimados por otras técnicas. Su mayor limitación estriba en la dificultad o imposibilidad de obtener un resultado que integre la heterogeneidad del suelo, de modo que generalmente, no se alcanza el tamaño de muestra preciso, y los resultados reflejan tanto, diferencias por concepto del lugar, como del tratamiento (Moreno 1994). A pesar de ello, son ampliamente empleados para estimar directamente el volumen de agua y nutrientes perdidos (Jordan 1982, Keller y Vedy 1991) que, aunque se reconocen sesgados, permiten obtener conclusiones, para fines de manejo.
Al Departamento de Ciencias Forestales de la Universidad Nacional de Colombia Sede Medellín, al Laboratorio de Ecología y Conservación Ambiental, adscrito al mismo Departamento, en particular a sus Laboratoristas Gloria Stella Mazo Carvajal y José Vicente Betancourt Ospina. Al Laboratorio del Área de Edafología y Química Agrícola de la Universidad de Salamanca, España y profesores asociados. Así mismo, al personal de la Escuela de Tecnología Forestal de Piedras Blancas por su apoyo en las tareas de campo. A las Empresas Públicas de Medellín por su autorización para realizar la investigación en predios de su propiedad y suministro de información climatológica.
Arias, L. A; González, L. H; Zapata, R.; Loaiza, J. C. y Arias, G. 2002. Comportamientos y estructuras del relieve y los suelos en el altiplano de Santa Rosa de Osos (Antioquia). Medellín: Universidad Nacional de Colombia, Sede Medellín. Facultad de Ciencias. Instituto de Ciencias Naturales y Ecología. Corporación Autónoma Regional del Centro de Antioquia - Corantioquia. 365 p. [ Links ]
Barreto, L. H. y León, J. D. 2005. Masa total y contenido de nutrientes en raíces finas de ecosistemas forestales (Pinus patula Schltdl y Cham, Cupressus lusitanica Mill y Quercus humboldtii, Bonpl.) en Piedras Blancas, Antioquia, Colombia. En: Revista Facultad Nacional de Agronomía Medellín. Vol. 58, no. 2; p. 2907-2929. [ Links ]
Baver, L. D.; Gardner, W. H. y Gardner, W. R. 1973. Física de Suelos. México: Hispano Americana. 529 p. [ Links ]
Beven, K. and German, P., 1981. Water flow in soil macropores, 2. A combined flow model. En: Journal of Soil Science. Vol. 32, no. 1; p. 15-29. [ Links ]
Bruijnzeel, L. A. 1984. Immobilization of nutrients in plantation forests of Pinus merkusii and Agathis dammara growing on volcanic soils in central Java, Indonesia . pp 19-29. En: Thajib, A. P., ed. Nutrition of Perrenial Crops. Kuala Lumpur: Malaysian Soil Science Society. 453 p. [ Links ]
________. 1991. Nutrient input-output budgets of tropical forest ecosystems: A review. En: Journal of Tropical Ecology. Vol. 7; p. 1-24. [ Links ]
________; Waterloo, M. J.; Proctor, J.; Kuiters, A. T. and Kotterink, B. 1993. Hydrological observations in montane rain forests on Gunung Silam, Sabah, Malaysia , with special reference to the Massenerhebung effect. En: Journal of Ecology. Vol. 81; p.145-167. [ Links ]
Cuevas, E. and Medina, E. 1988. Nutrient dynamics within amazonian forests. Part II. Fine root growth, nutrient availability and leaf litter decomposition. Oecologia, 76: 222-235. [ Links ]
Fassbender, Hans W. and Bornemisza, Elmer. 1987. Química de suelos con énfasis en suelos de América Latina. 2ed. San José de Costa Rica: Instituto Interamericano de Cooperación para la Agricultura. 420 p. [ Links ]
Food and Agriculture Organization of The United Nations. 2006. A framework for international classification, correlation and communication. Roma: FAO, ISRI and International Union of Soil Sciences. 128 p. (World Soil Resources Report 103). [ Links ]
Gavande, S. A. 1972. Física de suelos, principios y aplicaciones. México Limusa-Wiley. 351 p. [ Links ]
Gómez-Rey, M. X. y De Anta, C. 2002. Datos para el desarrollo de una red integrada de seguimiento de la calidad de suelos en Galicia (N.O de España): balances geoquímicos en suelos forestales (Pinus radiata). 2. Salidas de elementos y balance del suelo forestal. En: Edafología. Vol. 9, no. 2; p. 197-212. [ Links ]
Haines, B. L.; Waide, J. B. and Todd, R. L. 1982. Soil solution nutrient concentrations sampled with zero-tension lysi-meters: report of descrepancies. En: Soil Science Society of America Journal. Vol. 46; p. 658-661. [ Links ]
Herrera, R. 1979. Nutrient distribution and cycling in Amazon Caatinga forest on Spodosols in Southern Venezuela. Reading, U.K. 245 p. Thesis (Ph.D.), University of Reading. Department of Soil Science. [ Links ]
Herrera, R. A.; Jordan, C. F.; Klinge, H. and Medina, E. 1978. Amazon ecosystems: their structure and functioning with particular emphasis on nutrients. En: Interciencia. Vol. 3; p. 223-232. [ Links ]
Hillel, D. 1998. Environmental soil physics. San Diego, CA: Academic Press. 771 p. [ Links ]
Holdridge, L. R. 1987. Ecología basada en zonas de vida. San José, Costa Rica: Instituto Interamericano de Cooperación para la Agricultura. 216 p. [ Links ]
Huttel, C. H. 1975. Recherches sur l`écosystème de la forêt subèquatoriale de basse Côte d´Ivoire. IV Estimation du bilan hydrique. En:La Terre et la Vie. Vol. 29 ; p. 92-202. [ Links ]
Jaramillo, D. F. 1995. Andisoles del oriente antioqueño: caracterización química y fertilidad. Medellín: Universidad Nacional de Colombia. Facultad de Ciencias. 35 p. [ Links ]
________. 2002. Introducción a la ciencia del suelo. Medellín: Leo Digital. 53 p. [ Links ]
Jordan, C. F. 1969. Isotope cycles. En: The Rain Forests Project Annual Report. Puerto Rico Nuclear Center. p. 1-17. [ Links ]
________. 1982. The nutrient balance of an amazonian rain forest. En: Ecology. Vol. 63, no. 3; p. 647-654. [ Links ]
Kamprath, E. F. 1980. Soil acidity in welldrained soils of the tropics as a constraint to food production. p. 171-187. En: Soil- related constrains to food production in the tropics. Los Baños, Philippines : IRRI. [ Links ]
Keller, C. and Vedy, J. C. 1991. Apport de la lisimétrie sans tensions paur létude des transferts de Cu et Cd dans le sols forestiers faiblement pollués. En : Science du Sol. Vol. 29; p. 107-124. [ Links ]
Kenworthy, J. B. 1971. Water and nutrient cycling in a tropical rain forest. p. 49-65. En: Transactions of the First Aberdeen-Hull Symposium of Malesian Ecology. University of Hull, Department of Geography. 97 p. (Miscellaneous Series 11). [ Links ]
Lewis, W. M. Jr. 1986. Nitrogen and phosphorus runoff losses from a nutrient poor tropical moist forest. En: Ecology. Vol. 67; p. 275-1282. [ Links ]
Lilienfein, J.; Wilcke, W.; Ayarza, M. A.; Vilela, L.; Lima, S. D. C. and Zech, W. 2000. Soil acidification in Pinus caribaea forests on Brazilian savanna Oxisols. En: Forest Ecology and Management. Vol. 128; p. 145-157. [ Links ]
Londoño, A.; Montoya, D. C.; León, J. D. y González, M. I. 2005. Estudio del flujo saturado de agua en suelos forestales de Piedras Blancas, Antioquia. En: Boletín Ciencias de la Tierra. Vol. 17; p. 73-84. [ Links ]
Marques, R. and Ranger, J. 1997. Nutrient dynamics in a chronosequence of Douglas fir (Pseudotsuga menziesii (Mirb.) Franco) stands on the Beaujolais Mounts (France). Part 1. Qualitative approach. En: Forest Ecology and Management. Vol. 91; p. 255-277. [ Links ]
Moreno, M. G. 1994. Balances de agua y nutrientes en Rebollares (Quercus pyrenaica Willd.) de la vertiente Salmantina de la Sierra de Gata. Salamanca. 470 p. Tesis Doctoral. Universidad de Salamanca. [ Links ]
Moreno, G.; Gallardo, J. F.; Schneider, K. and Ingelmo, F. 1996. Water and bioelement fluxes in four Quercus pyrenaica forest along a pluviometric gradient. En: Annales des Sciences Forestières. Vol. 53; p. 625-639. [ Links ]
Nilsson, L. O.; Hüttl R. F.; Johansson, U. T. and Jochheim, H. 1995. Nutrient uptake and cycling in forest ecosystems-present status and future research directions. En: Plant and Soil. Vol. 168-169, no.1; p. 5-13. [ Links ]
Parker, G. G. 1985. The effect of disturbance on water and solute budgets of hills slope tropical rainforest in Northeastern Costa Rica. Athens, GA. 161 p. Thesis. (Ph.D.) University of Georgia. [ Links ]
Ranger, J.; Marques, R. and Jussy, J-H. 2001. Forest soil dynamics during stand development assessed by lysi-meter and centrifuge solutions. En: Forest Ecology and Management. Vol. 144, no. 1-3; p. 129-145. [ Links ]
Ruiz S., O. A.; Acosta, J. J. y León P., J. D. 2005. Escorrentía superficial en bosques montanos naturales y plantados de Piedras Blancas, Antioquia ( Colombia). En: Revista Facultad Nacional de Agronomía, Medellín. Vol. 58, no.1; p. 2635-2649. [ Links ]
Russell, C. E. 1983. Nutrient cycling and productivity of native and plantation forests of Jari Florestal, Pará, Brazil . Athens, GA. 130 p. Thesis. (Ph.D.) University of Georgia. [ Links ]
Soil Survey Staff. 2003. Keys to soil taxonomy. 9 ed. Washington: United States Department of Agriculture. Natural Resources Conservation Service. 332 p. [ Links ]
Tschinkel, H. 1972. Factores limitantes del crecimiento de plantaciones de Cupressus lusitanica en Antioquia, Colombia. En: Revista Facultad Nacional de Agronomía, Medellín. Vol. 27, no. 2; p. 3-55. [ Links ]
Turvey, N. D. 1974. Nutrient cycling under tropical rain forest in Central Papua. New Guinea: Department of Geography, University of Papua. 96 p. (Occasional paper, N 10). [ Links ]
Universidad Nacional de Colombia. 1999. Cuenta del bosque cuenca de Piedras Blancas (Antioquia). Medellín: La Universidad. 96 p. [ Links ]
Vitousek, P. M. and Sanford, R. L. 1986. Nutrient cycling in moist tropical forest. En: Annual Review of Ecology and Systematics. Vol. 17; p. 137-167. [ Links ]
Voigt, G. K. 1980. Acid precipitation and soil buffering capacity: p. 53-57. En: Drablos, D, and Tollan, A., eds. Proceedings of the International Conference on the Ecological Impact of Acid Precipitation. Sandefjord, Norway . [ Links ]
Zabowski, D. and Ugolini, F. C. 1990. Lysimeters and centrifuge soil solutions: seasonal differences between methods. Soil Science Society of America Journal. Vol. 54, no.1; p. 1130-1135. [ Links ]
Zech, W., Wilcke, W. and Valarezo, C. 1999. Influencia del uso de la tierra en las propiedades del suelo en los flujos de agua y de elementos en los bosques montañosos del Ecuador del sur. Investigación Científica del Instituto Ecuatoriano Forestal de Áreas Naturales y Vida Silvestre (INEFAN). Quito, Ecuador. 57 p. [ Links ]

