










Services on Demand
Journal
Article
 Spanish (pdf)
Spanish (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mailIndicators
-
 Cited by SciELO
Cited by SciELO -
 Access statistics
Access statistics
Related links
-
 Cited by Google
Cited by Google -
 Similars in
SciELO
Similars in
SciELO -
 Similars in Google
Similars in Google
Share

 Permalink
PermalinkRevista Facultad Nacional de Agronomía Medellín
Print version ISSN 0304-2847
Rev. Fac. Nac. Agron. Medellín vol.63 no.2 Medellín July/Dec. 2010
RELACIÓN ENTRE PÉRDIDA DE PESO, PERFIL LIPÍDICO Y CONCENTRACIONES PLASMÁTICAS DE LEPTINA EN VACAS CEBÚ PRIMERIZAS
RELATIONSHIP AMONG WEIGHT LOSS, LIPIDIC PROFILE AND PLASMA CONCENTRATIONS OF LEPTIN IN FIRST CALVING ZEBU COWS
Guillermo Henao Restrepo1; Rubén Darío Galvis Goez2; Liliana María Cardona Bermúdez3 y Natalia María Castro Henao4
1 Profesor Titular. Universidad Nacional de Colombia, Sede Medellín. Facultad de Ciencias Agropecuarias. A.A.1779, Medellín, Colombia. <ghenao@unal.edu.co>
2 Profesor Asociado. Universidad Nacional de Colombia, Sede Medellín. Facultad de Ciencias Agropecuarias. A.A.1779, Medellín, Colombia. <rdgalvis@unal.edu.co>
3 Zootecnista. Universidad Nacional de Colombia, Sede Medellín. Facultad de Ciencias Agropecuarias. A.A.1779, Medellín, Colombia. <lilichurra@gmail.com>
4 Zootecnista. Universidad Nacional de Colombia, Sede Medellín. Facultad de Ciencias Agropecuarias. A.A.1779, Medellín, Colombia. <nmcastroh@gmail.com>
Recibido: Septiembre 4 de 2009; Aceptado: Mayo 4 de 2010
Resumen. Para caracterizar las relaciones existentes entre la pérdida de peso, el perfil lipídico y la concentración de leptina durante el posparto, se seleccionaron 20 novillas Cebú sanas con 8,2±0,6 meses de gestación y condición corporal entre 6 y 8 (escala 1-9). Cada 14 días se midió el peso y semanalmente se tomaron muestras de sangre desde el ingreso al experimento hasta 56 días posparto. Se determinaron y compararon los niveles de ácidos grasos no esterificados (AGNES), colesterol, leptina y perfil de lipoproteínas (HDL, LDL, VLDL); se evaluaron las relaciones entre los cambios de peso y el perfil lipídico y entre los cambios de peso y la leptina plasmática. Respecto a los cambios de peso se presentaron diferencias (P<0,05) entre las semanas posparto, sugiriendo una menor disminución en la proporción de pérdida de peso a medida que avanzaba el periodo posparto. En cuanto al efecto de los días posparto sobre el perfil lipídico, se apreciaron variaciones significativas para colesterol, lipoproteínas y AGNES. Se encontró relación significativa y directa entre colesterol y leptina, y entre la pérdida de peso y los niveles de AGNES. Se registraron relaciones significativas e inversas de AGNES con colesterol y con HDL. Lo anterior sugiere que en el posparto temprano las vacas Cebú primerizas en un principio, presentan valores bajos de LDL y posteriormente de HDL, sin ningún cambio en los de VLDL, lo que puede limitar la adecuada movilización y distribución de las reservas corporales hasta el hígado y desde éste a los demás órganos.
Palabras clave: Agnes, lipoproteínas, movilización lipídica, posparto
Abstract. To characterize the existing relations among weight loss, lipid profile and generation of leptin during the postpartum period, 20 healthy Zebu heifers with 8,2 ±0,6 months of gestation and body condition score between 6 and 8 (1-9 points scale) were selected. Every 14 days the cow’s live weight was measured and weekly blood samples were taken from the start of the experiment up to 56 days postpartum. The levels of non-esterified fatty acids (NEFAS), cholesterol, leptin and lipoproteins profile (HDL, LDL, VLDL), were quantified, evaluated and compared; the possible relationships among cow live weight change with lipid profile and plasmatic leptin were evaluated. Respect to the changes of weight, differences (P<0.05) between weeks postpartum appeared, suggesting a smaller diminution in the proportion of loss of weight as advanced the postpartum period. Respect to the effect of the days postpartum on the lipid profile, significant variations for cholesterol, lipoproteins and NEFAS appeared. There were significant and direct relation between cholesterol and leptin, and the loss of weight and NEFAS. There were significant and inverse relations of NEFAS with cholesterol and HDL. All of the above suggests that in the early postpartum, at the beginning the primiparous Zebu cows show low values of LDL, and HDL later, without any change in the VLDL, which may limit the proper mobilization and distribution of body reserves to the liver and from it to other organs.
Key words: Nefas, cholesterol, lipoproteins, postpartum.
Debido a las condiciones climáticas adversas de las zonas tropicales, se dificulta desarrollar sistemas productivos eficientes con bovinos Bos taurus, por lo cual los ganaderos recurren con más frecuencia a la producción con bovinos Bos indicus, siendo las distintas razas de bovinos Cebú las más distribuidas en estas zonas debido a su gran rusticidad. A pesar de lo anterior, las condiciones nutricionales predominantes en las zonas tropicales inducen aceleradas pérdidas de peso y de condición corporal que se dan después del parto y que simultáneamente alteran el estado metabólico del animal e indirectamente la reproducción. Tradicionalmente la producción con estas razas se realiza en condiciones de pastoreo extensivo con amamantamiento permanente del becerro.
Las condiciones tropicales someten a estos animales a una disponibilidad y calidad variable de los pastos durante el año (FEDEGAN, 2006), los cuales son más abundantes y nutritivos en períodos con adecuada distribución de las lluvias en los que las vacas los consumen a voluntad, aumentan peso y condición corporal, desarrollan ciclos estrales y conciben. Durante los períodos de sequía los pastos son escasos y fibrosos, las vacas bajan el consumo de alimento, disminuyen peso y reservas corporales y desarrollan un anestro prolongado que afecta negativamente la eficiencia reproductiva (Montiel y Ahuja, 2005).
Los diferentes grados de subnutrición derivados de un bajo consumo de pastos con respecto a los requerimientos nutricionales del animal, producen cambios en la condición corporal de las vacas, por movilización de reservas energéticas que ocasionan cambios en algunos metabolitos indicadores del estado energético (Herdt, 2000). Las posibles causas de este fenómeno no sólo son debidas a factores medioambientales como el clima o la disponibilidad de alimentos, sino también a factores propios del animal, como los asociados con la menor capacidad de consumo de forrajes y de respuesta metabólica ante las demandas del parto y la producción de leche (Galvis et al, 2003).
La baja ingestión de materia seca digestible conduce a balances energéticos (BEN) y proteicos negativos que se hacen más pronunciados en el posparto temprano. Ante las condiciones nutricionales deficientes la respuesta metabólica del animal induce la movilización de reservas corporales para suplir el déficit, lo cual conlleva a pérdidas de peso y cambios significativos en la condición corporal (Herdt, 2000).
La movilización de reservas corporales es un mecanismo que permite la exportación de sustratos lipídicos energéticos desde el tejido adiposo a la circulación sanguínea. De esta manera salen de los adipocitos grandes cantidades de ácidos grasos no esterificados (AGNES) con el objetivo de ser oxidados en la mayoría de los tejidos. El hígado es uno de los principales órganos en utilizar los AGNES, además de ser el nodo central desde donde se redistribuye el flujo de lípidos hacia todo el organismo. En el hígado se ensamblan continuamente lipoproteínas de muy baja densidad (VLDL), con el único objetivo de distribuir las fracciones lipídicas desde este órgano a los demás; de igual manera una intensa síntesis de triglicéridos es llevada allí (Kaneko, 1989); por consiguiente, la capacidad de movilización de fracciones lipídicas desde el hígado es muy importante para redistribuir las reservas de energía y para impedir la acumulación de grasa en dicho órgano, situación probable en vacas sometidas a BEN pronunciados (Galvis et al., 2003).
Se ha postulado una relación directa entre el nivel de reservas corporales y la hormona leptina (Strauch et al., 2003). Esta es una hormona producida por los adipocitos, cuya función principal es regular la ingesta y la termogénesis. Es probable que en las vacas con BEN haya una deficiencia en la secreción de leptina desde el tejido adiposo, lo cual alteraría el consumo voluntario de forraje y con esto la dinámica normal del balance energético. Una vez que se recupera el equilibrio energético, la leptina tiene profundos efectos sobre la modulación de la distribución de reservas y de la iniciación de la actividad reproductiva (Recabarren, 2003).
El objetivo de este trabajo fue explorar las relaciones existentes entre la pérdida de peso corporal, el perfil lipídico y la generación de leptina en vacas Cebú primerizas en amamantamiento durante el posparto temprano.
MATERIALES Y MÉTODOS
Esta investigación se realizó en las haciendas Buenos Aires y La Sarana ubicadas en el municipio de Cimitarra, departamento de Santander (Colombia), en una zona de vida de bosque húmedo tropical (bh-T) (Holdridge, 1978) a 200 msnm, con precipitación de 3.833 mm y temperatura de 30 °C. La evaluación de niveles de metabolitos y hormonas se realizó en los laboratorios AGROLAB y de Fisiología Animal de la Universidad Nacional de Colombia, Sede Medellín.
Animales y diseño experimental. Se seleccionaron 20 novillas Cebú con 8,2 ± 0,6 meses de gestación, con buen estado de salud y una condición corporal entre 6 y 8 (escala 1 a 9 puntos) (Hardin, 1990). Todos los animales pastorearon praderas de Brachiaria decumbens (6,5% de proteína) y B. humidicola (7,2% de proteína) en un sistema rotacional con una oferta promedio diaria de 5,5% de su peso corporal en materia seca, estimada por la técnica de botanal (Giraldo, 1996) el día anterior a la ocupación de cada potrero. Las novillas fueron sometidas durante dos semanas a un programa de amanse y acostumbramiento al muestreo.
Determinación del perfil lipídico y concentración de leptina. El día de la selección, el día del parto y cada semana hasta los 56 días posparto se tomaron muestras de 10 mL de sangre en tubos al vacío, por punción de los vasos coccígeos. Las muestras se dejaron a temperatura ambiente por un tiempo aproximado de tres horas, se centrifugaron a 1800 x g durante cinco minutos y el suero se conservó en cinco alícuotas en nitrógeno líquido a -196 °C hasta la cuantificación de AGNES, colesterol total, leptina, triglicéridos y lipoproteína de alta densidad (HDL). Se midieron las concentraciones circulantes de triglicéridos, colesterol total, y HDL utilizando los kits comerciales BioSystems®. La estimación de lipoproteína de muy baja densidad (VLDL) se evaluó por diferencia según la propuesta de Friedewald (Working Group on Lipoprotein Measurement, 1995). La precisión de cada determinación se monitoreó usando una muestra de suero comercial de referencia. Las concentraciones séricas de leptina se determinaron por radioinmunoanálisis utilizando el kit comercial XL-85-K® (LINCO, Missouri, USA) y las concentraciones séricas de AGNES se precisaron utilizando el kit comercial de ensayo enzimático acoplado a espectrofotometría AGNE-C® 994-75409 E (Wako, Dallas TX).
Determinación del peso y la condición corporal. El peso corporal de cada novilla se midió al momento de la selección, el día del parto y luego cada periodo de 14 días hasta los 56 días posparto, utilizando una báscula ganadera con escala de 1 a 1500 kg, calibrada antes del pesaje de cada animal. Debido a que fue imposible determinar la pérdida de peso de la madre al momento del parto, sin que el peso del feto, las membranas y líquidos fetales interfirieran en su precisión, se optó por analizar solo los cambios de peso durante el período posparto. Con un intervalo de 15 días se evaluaron los cambios de peso entre el parto y la semana 8 posparto, este se reportó como porcentaje respecto al peso vivo al momento del parto. Por su parte la condición corporal se evaluó en una escala de 1 a 9 siguiendo la metodología propuesta por Dieter, Diaz y Florez (1999).
Análisis estadístico. Se evaluaron los efectos de los días posparto sobre los cambios de peso y sobre las concentraciones plasmáticas de AGNES, colesterol, leptina, VLDL, Triglicéridos, LDL y HDL por medio de un análisis de varianza usando el PROG GLM del paquete estadístico SAS® (SAS Institute Inc, 1988). Se evaluaron las posibles relaciones entre los cambios de peso y concentración de metabolitos y leptina mediante análisis de regresión PROC REG de SAS® (SAS Institute Inc, 1988). Se aceptaron diferencias estadísticamente significativas cuando P<0,05.
RESULTADOS
Perfil lipídico y leptina. Se presentaron variaciones en las concentraciones de colesterol (P<0,05), HDL (P<0,05), LDL (P<0,05) y AGNES (P<0,05), dependientes de los días posparto. Las concentraciones de leptina, VLDL y triglicéridos no presentaron variaciones significativas entre los periodos de muestreo. Las Figuras 1 y 2 ilustran la evolución del perfil lipídico, de los AGNES y de la leptina durante los periodos de muestreo.
Cambios de peso y condición corporal. Se presentaron diferencias (P<0.05) para las pérdidas de peso y condición corporal, lo que sugiere una disminución en la tasa de pérdida de peso conforme avanzaron los días posparto. Los cambios de peso y condición corporal se pueden observar en la Figura 3.
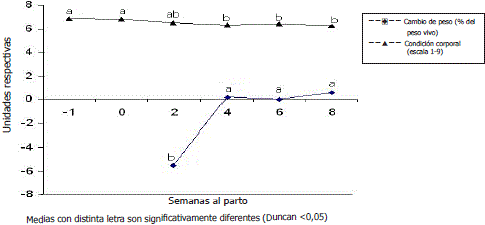
Figura 3. Evolución del cambio de peso y la condición corporal en vacas Cebú primerizas.
Asociación entre variables. En la Tabla 1 se consignan las asociaciones exploradas entre las diferentes variables. Se encontraron relaciones directas entre las concentraciones séricas de colesterol y leptina (P<0,05) (Figura 4); una relación inversa entre el cambio de peso y las concentraciones séricas de AGNES (P<0,05) (Figura 5). De igual manera se encontraron relaciones inversas entre las concentraciones séricas de AGNES y colesterol (P<0,05) (Figura 6) y entre las concentraciones séricas de AGNES y de HDL (P<0,05) (Figura 7). No se presentaron relaciones significativas entre los cambios de peso y las concentraciones séricas de leptina y entre las concentraciones séricas de AGNES y las de triglicéridos, VLDL y leptina (Tabla 1).
Tabla 1. Ecuaciones de regresión entre las diferentes variables consideradas para estudiar las variaciones fisiológicas posparto en vacas Cebú primerizas.
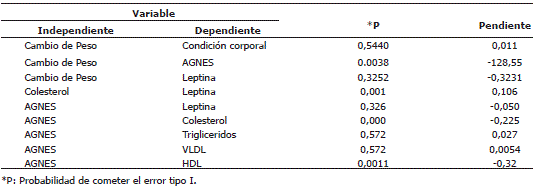

Figura 4. Concentraciones de leptina en relación con las de colesterol en vacas Cebú primerizas.

Figura 5. Concentraciones de AGNES con relación a los cambios de peso en vacas Cebú primerizas.

Figura 6. Concentraciones de colesterol en relación con las de AGNES en vacas Cebú primerizas.
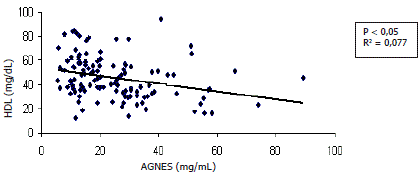
Figura 7. Concentraciones de HDL en relación con las de AGNES en vacas Cebú primerizas
DISCUSIÓN
Durante el periodo posparto las vacas perdieron peso debido posiblemente a que después del parto tienen una menor capacidad de consumo de forrajes, lo que les imposibilita cubrir las demandas que trae consigo la producción de leche. La baja ingestión de materia seca digestible conduce a balances energéticos (BEN) y proteicos negativos que se hacen más pronunciados en el posparto temprano. Ante las condiciones nutricionales deficientes la respuesta metabólica de las vacas induce la movilización de reservas corporales para suplir el déficit; esto conlleva a pérdidas de peso y cambios significativos en la condición corporal (Herdt, 2000). Se pudo evidenciar que a medida que avanzaban los días en lactancia disminuía la pérdida de peso corporal, como consecuencia de una disminución en los requerimientos de nutrientes que conduce a una recuperación del peso y a una normalización del metabolismo energético del animal, lo que concuerda con los hallazgos de Galvis et al. (2003) en ganado lechero, sin embargo, este fenómeno se presentó más temprano en las vacas Cebú primerizas que en vacas lecheras. También se observó que a medida que aumentaban los días en lactancia, disminuía la condición corporal, de manera similar al hallazgo de Ceballos y Andaur (1999) en vacas lecheras, quienes afirman que después del parto se presenta una pérdida acentuada de la condición corporal.
Respecto al nivel plasmático de colesterol se encontró un efecto significativo de los días posparto. De esta forma, el nivel de colesterol aumentó a medida que avanzaron los días posparto; lo anterior concuerda con los resultados encontrados por varios investigadores (Aranda, Brave y Casagrande (2003); Galvis et al. (2003) quienes afirman que las concentraciones de colesterol son bajas en el periparto y aumentan a través del posparto, lo cual es consecuencia de una disminución en los requerimientos de nutrientes y de una normalización del metabolismo energético de las vacas. Estos resultados reafirman lo enunciado por Ceballos et al. (2002), Galvis et al. (2003), Galvis, Agudelo y Saffon (2007), quienes expresan que la concentración de colesterol, es un indicador confiable del balance de energía desde un mes preparto hasta la semana seis posparto; por su parte, Ceballos et al. (2002), agregan que a la sexta semana posparto la vaca, bien sea de baja o alta producción, alcanza su equilibrio metabólico después del parto, lo que se ve reflejado en el nivel de colesterol plasmático. Según estas aseveraciones es lógico pensar que en vacas Bos indicus el nivel de colesterol plasmático es un buen indicador del estado nutricional y metabólico.
Los triglicéridos plasmáticos son un indicador de la disponibilidad energética del animal (Kaneko, 1989), el cual realiza una síntesis neta de éstos, solo cuando la disponibilidad de energía lo permite, por eso la concentración de triglicéridos plasmáticos aumenta conforme disminuye el déficit energético (Grummer, 1993) a medida que avanza la lactancia (West, Multinix y Sandifer, 1991); sin embargo, en esta investigación las concentraciones plasmáticas de triglicéridos no aumentaron significativamente, a través del periodo posparto, lo que pudo ser debido al estado nutricional y a la dinámica del metabolismo particular de las vacas Bos indicus, las cuales no producen altas cantidades de leche durante el posparto temprano, lo que probablemente origina cambios metabólicos menos evidentes a los observados en vacas productoras de leche.
En el presente estudio las vacas no incrementaron las concentraciones plasmáticas de VLDL conforme avanzó la lactancia, lo que pudo ser debido a la baja capacidad de síntesis en el hígado observada en rumiantes (Kleppe et al, 1988; Pechová, Illek y Halouzca,1997), principalmente en la lactancia temprana (Herdt, 2000). Según varios autores (Grummer, 1993; Pechová, Illek y Halouzca, 1997 y Van Den Top et al. 1996) el aumento en los valores plasmáticos de VLDL ocurre cuando el animal se acerca a un balance energético positivo como consecuencia de un mayor consumo de nutrientes y de menores gastos energéticos para la producción, esto concuerda con lo observado en esta investigación, dado que las vacas tuvieron pérdidas moderadas de peso durante todo el periodo posparto lo que sugiere una condición energética desfavorable durante todo este periodo. Respecto a la lipoproteína de alta densidad (HDL), hubo un incremento en su concentración a través del periodo posparto, lo que es coherente con Bauchart (1993) y Basoglu, Sevinc y Gokcen, (1998); quienes afirman que la HDL aumenta a medida que avanzan los días en lactancia y que el valor máximo se alcanza en la lactancia tardía. Sin embargo, Rayssiguier et al. (1988), citados por Ceballos et al. (2002); encontraron que después del parto la concentración de HDL aumentó hasta alcanzar valores elevados cuatro semanas posparto. Según Puppione (1978), durante la lactancia temprana el metabolismo acelerado de la VLDL origina la subsiguiente cobertura de lípidos remanentes en la superficie capilar, que posteriormente formará las HDL y las cuales probablemente tengan una baja tasa de recambio. Este fenómeno da cuenta de las elevadas concentraciones de HDL durante este período. Al respecto, Kaneko (1989) sugiere tres posibles explicaciones para los niveles altos de HDL en el plasma de vacas en lactación: uno es la adaptación a la lactancia incrementando el reservorio de apoproteína C, otro es la utilización incrementada de VLDL por la glándula mamaria y por último síntesis y secreción de HDL naciente, incrementada por el hígado en respuesta a la lactación.
Dado que no se evidenciaron aumentos en las concentraciones de VLDL, es de esperar que la elevación en las concentraciones de HDL, no estuviera asociado al incremento en la utilización de VLDL por la glándula mamaria. Las concentraciones plasmáticas de LDL crecieron conforme avanzó la lactancia; ello concuerda con reportes de Bauchart (1993), quien afirma que los receptores para LDL aumentan conforme aumentan los niveles de estrógenos, por lo tanto, debido a que durante la lactancia temprana la secreción de estrógenos ováricos es escasa, hay una disminución en los receptores y por ende hay mayor concentración de LDL circulante. Es posible además que un aumento en las concentraciones de colesterol por efecto de los días posparto conlleve al incremento en esta lipoproteína, necesaria para su transporte y distribución (Barahona, Blume y Bruno, 2005). Una posibilidad adicional para la elevación de la LDL, consiste en el incremento de VLDL por los tejidos periféricos (Kaneko, 1989), entre ellos la glándula mamaría; de esta manera es de esperar que en las semanas previas al pico de producción de leche se incrementen las concentraciones de LDL, debido a un aumento en la captación de VLDL por la glándula mamaria.
Las concentraciones de AGNES variaron significativamente acorde con los días posparto. Se pudo evidenciar que la concentración de AGNES aumentó durante las dos primeras semanas posparto y posteriormente disminuyó. Lo que es afín con lo expuesto por Herdt (2000) quien postula que en estado de BEN, grandes cantidades de AGNES son movilizadas del tejido adiposo para ser utilizados como fuente de energía por los demás tejidos. Adicionalmente, la hormona somatotropina tiene un papel importante en la regulación del uso de los combustibles en el BEN; ésta reduce la lipogénesis favoreciendo la movilización de AGNES, lo que cobra mayor magnitud en la lactancia temprana, dado que la concentración de somatotropina se encuentra aumentada en este periodo (West, Multinix y Sandifer, 1991). Una posible explicación para la disminución de las concentraciones de AGNES, es la merma de los requerimientos energéticos conforme se rebaja la producción de leche, lo que se traduce teóricamente en un aumento en la disponibilidad de glucosa. Herdt (2000) en una amplia revisión sugiere que cuando aumenta la disponibilidad de glucosa, aumenta la tasa de lipogénesis respecto a la de la lipólisis en el adipocito; esto conlleva a una baja liberación de AGNES desde el tejido adiposo y por consiguiente disminuyen la concentración de AGNES en la sangre.
Las concentraciones de leptina no presentaron variaciones significativas a través de los días posparto, fenómeno no esperado ya que a medida que las vacas se acercaban al momento de máxima pérdida de peso, debería ocurrir una disminución en la concentraciones plasmáticas de leptina. Recabarren (2003) sugiere que es probable que en las vacas en estado de BEN haya una deficiencia en la secreción de leptina desde el tejido adiposo. Por su parte Delavaud et al. (2002) han encontrado en sus estudios efectos significativos del nivel de alimentación sobre las concentraciones de leptina, donde ésta aumenta en la medida en que las vacas reciben una dieta que llena completamente sus requerimientos y también en aquellas que son sobrealimentadas. Adicionalmente estos mismos autores afirman que la concentración de leptina en plasma depende principalmente del tamaño de la célula del adipocito más que de la condición corporal, efecto éste que puede observarse a largo plazo, pero que no fue evaluado en la presente investigación.
Asociación entre variables. El cambio de peso se relacionó negativamente con la concentración de AGNES, la cual depende de la cantidad de reservas corporales que el animal moviliza en momentos en los que los nutrientes suministrados en la dieta no son suficientes para suplir los requerimientos. Como ya se mencionó (Herdt, 2000) en estado de BEN grandes cantidades de AGNES son movilizadas del tejido adiposo para ser utilizados como fuente de energía por los demás tejidos, por lo tanto es de esperarse una relación directa entre las pérdidas de peso y las concentraciones plasmáticas de AGNES.
Se encontró una relación directa entre las concentraciones plasmáticas de leptina y colesterol, lo cual coincide con los hallazgos de Hamman y Matthaei (1996), quienes afirman que la producción de leptina aumenta conforme aumenta el contenido adiposo del cuerpo y el suministro de nutrientes, ya que cuando se inicia la recuperación de la condición corporal a través del tiempo, se produce un incremento en los niveles de colesterol. Galvis, Agudelo y Saffon (2007) encontraron una correlación marcada de la secreción de leptina con el contenido adiposo corporal y con el tamaño de los adipocitos; sin embargo, estos mismos autores afirman que su correlación con el tejido adiposo se ve fuertemente alterada por cambios en el balance energético y por la privación de alimento (por un tiempo de 12 a 48 horas) lo que resulta en una rápida y dramática caída en la expresión del gen de la leptina; cambios sutiles en el balance energético tienen igualmente profundos efectos en su expresión. Aún no está claro si los efectos de la condición corporal sobre los niveles plasmáticos de leptina se ven alterados por el número y tamaño de los adipocitos; es así como Delavaud et al. (2002), encontraron que las concentraciones plasmáticas de leptina se relacionan más directamente con el tamaño de las células adiposas que con la condición corporal; parámetro que no se determinó en esta investigación.
Se encontró una relación inversa entre las concentraciones plasmáticas de AGNES y colesterol. Aranda, Brave y Casagrande (2003) establecieron que la concentración sérica de AGNES es alta durante el periparto mientras que la de colesterol está disminuida, corroborando de esta manera el tipo de asociación encontrada en el presente estudio. Como se indicó antes, el colesterol aumenta conforme se recupera el estatus energético del animal, situación que conlleva a su vez a que se disminuya la movilización de reservas corporales y con ello la concentración de AGNES en el plasma sanguíneo. Por su parte Sommer (1995); citado por Ceballos et al. (2002), confirma esta asociación, manifestando que la colesterolemia constituye un reflejo del consumo de energía de la ración.
Los valores plasmáticos de LDL aumentaron conforme disminuyó la condición corporal de la vaca y ésta a su vez se redujo a medida que avanzó la lactancia. Bauchart (1993) afirma que conforme avanza la lactancia se presenta una disminución en los receptores para LDL como consecuencia de la disminución de los estrógenos; de esta forma es probable que en la presente investigación el aumento de las concentraciones plasmáticas de LDL, asociado inversamente con la condición corporal, sea un efecto indirecto en la disminución de sus receptores, más que una asociación biológica entre estas variables.
Hubo un aumento de HDL conforme se redujeron los valores plasmáticos de AGNES, porque cuando el animal realiza exportación de lípidos desde el hígado hacia los demás tejidos, lo hace en forma de VLDL, que según Puppione (1978), origina la subsiguiente cobertura de lípidos remanentes en la superficie capilar que posteriormente formará las HDL, probablemente con una baja tasa de recambio. Por lo tanto, la elevación de los niveles de HDL debe estar asociada a estados de lipogénesis en vez de lipólisis, del cual los AGNES son su mejor indicador. De igual forma Aranda, Brave y Casagrande (2003), han reportado que en general, la concentración de los ácidos grasos no esterificados está aumentada en el periparto, mientras que la de colesterol está disminuida; probablemente en todas sus fracciones incluyendo la HDL.
En conclusión, se presentaron diferencias en cuanto a cambios de peso, condición corporal y AGNES entre las semanas posparto, sugiriendo una disminución en la pérdida de peso a medida que avanzaba el posparto. En cuanto al efecto de los días posparto sobre el perfil lipídico, se presentaron aumentos significativos en colesterol, HDL y LDL conforme avanzaba la lactancia y disminuían las pérdidas de peso, denotando una condición energética más favorable. Por el contrario, no se encontraron variaciones significativas en las concentraciones plasmáticas de triglicéridos, VLDL y leptina. En términos generales se pudo evidenciar una sincronía entre los valores de colesterol y de leptina, ambos metabolitos reconocidos como indicadores de situación nutricional favorable. No obstante se encontraron relaciones inversas de AGNES con colesterol y con HDL, sugiriendo que las vacas Cebú primerizas en el posparto temprano presentan en un principio bajos valores de LDL y posteriormente de HDL, sin ningún cambio en los de VLDL, lo que puede limitar la adecuada movilización y distribución de las reservas corporales hasta el hígado y desde éste a los demás órganos deficitarios en energía.
AGRADECIMIENTOS
A la Dirección de Investigaciones de la Universidad Nacional de Colombia, Sede Medellín (código DIME: 030803756), por la financiación de ésta investigación. A las haciendas Buenos Aires y La Sarana. A Gloria Fonegra, laboratorista Agrolab Ltda. Al profesor Héctor Jairo Correa. A Jorge Gómez, Profesor Politécnico Jaime Isaza Cadavid y a Mary Luz Jiménez, Ingeniera Agropecuaria.
BIBLIOGRAFÍA
Aranda, M.V., N. Brave y R. Casagrande. 2003. Colesterol en bovinos. En: Sitio argentino de producción animal, http://www.produccion-animal.com.ar/produccion_bovina_de_carne.htm; consulta: julio 2005. [ Links ]
Barahona, K., C. Blume y J.I. Bruno. 2005. Lipoproteínas sanguíneas: tipos e importancia. En: Seminario química biológica Universidad San Martín de Porres, http://www.monografias.com/trabajos16/lipoproteinas-sanguineas/lipoproteinas-sanguineas2.shtml.; consulta: julio 2005. [ Links ]
Basoglu, A., M. Sevinc and M. Gokcen. 1998. Peri- and postparturient concentrations of lipid lipoprotein, insulin and glucose in normal dairy cows. Tukish Journal of Veterinary and Animal Science 22: 141-144. [ Links ]
Bauchart, D. 1993. Lipid absorption and transport in rumiants. Journal of Dairy Science 76(12): 3864-3881. [ Links ]
Ceballos, M.A., R.M. Andaur. 1999. Indicadores bioquímicos sanguíneos de los desequilibrios energéticos en ganado lechero. p. 17-44. En: Memorias Seminario Internacional de Reproducción y Metabolismo de la Vaca Lechera. Universidad de Caldas, Manizales. [ Links ]
Ceballos, A., P.M. Gómez, M.L. Vélez, N.A. Villa y López, L.F. 2002. Variación de los indicadores bioquímicos del balance de energía según el estado productivo de los bovinos lecheros de Manizales, Colombia. Revista Colombiana de Ciencias Pecuarias 15(1): 13-25. [ Links ]
Delavaud, C., A. Ferlay, Y. Faulconnier, F. Bocquier, G. Kann and Y. Chilliard. 2002. Plasma leptin concentration in adult cattle: Effects of breed, adiposity, feeding level, and meal intake. Journal of Animal Science 80(5): 1317-1328. [ Links ]
Dieter H., T. Diaz y H. Florez. 1999. Evaluación de la condición corporal en vacas doble propósito. Manual Corpoica, Tibaitatá. 12 p. [ Links ]
Federación Colombiana de Ganaderos – FEDEGAN. 2006. Plan estratégico de la ganadería colombiana 2019. 1ª ed. Editorial San Martín Obregón, Bogotá, Colombia.
Galvis, R.D., H.J. Correa, N.F. Ramirez y W. Soler. 2003. Influencia de las alteraciones hepáticas sobre la actividad PEPCK, IGF-I plasmático y la reactivación ovárica en la lactancia temprana. Revista Colombiana de Ciencias Pecuarias 16(4): 228-236. [ Links ]
Galvis, R.D., D. Agudelo y A. Saffon. 2007. Condición corporal, perfil de lipoproteínas y actividad ovárica en vacas Holstein en lactancia temprana. Revista Colombiana de Ciencias Pecuarias 20(1): 16-29. [ Links ]
Giraldo, L. 1996. Manejo y utilización sostenible de pasturas. Universidad Nacional de Colombia, Medellín. 342 p. [ Links ]
Grummer, R.R. 1993. Etiology of lipid-related metabolic disorders in periparturient dairy cows. Journal of Dairy Science 76(12): 3882-3896. [ Links ]
Hamman, A. and S. Matthaei. 1996. Regulation of energy balance by leptin. Experimental and Clinical Endocrinology and Diabetes. 104(4): 293-300. [ Links ]
Hardin, R. 1990. Using body condition scoring in beef cattle management. University of Georgia College of Agricultural and Environmental Sciences, Cooperative Extension Service, C-817. 4 p. [ Links ]
Herdt, T.H. 2000. Ruminant adaptation to negative energy balance: influences on the etiology of ketosis and fatty liver. The Veterinary Clinics of North America. Food Animal Practice 16(2):215-230. [ Links ]
Holdridge, L.R. 1978. Ecología basada en zonas de vida. Instituto Interamericano de Ciencias Agrícolas, San José Costa Rica. 216 p. [ Links ]
Kaneko, J.J. 1989. Clinical Biochemistry of Domestic Animals. Department of Clinical Pathology, School of Veterinary Medecine, University of California. 930 p. [ Links ]
Kleppe, B.B., R.J. Aiello, R.R. Grummer and L.E. Armentano. 1988. Triglyceride accumulation and very low density lipoprotein secretion by rat and goat hepatocytes in vitro. Journal of Dairy Science 71(7):1813. [ Links ]
Montiel, F. and C. Ahuja. 2005. Body condition and suckling as factors influencing the duration of postpartum anestrus in cattle: a review. Animal Reproduction Science 85(1-2): 1-26. [ Links ]
Pechová, A., J. Illek and R. Halouzca. 1997. Diagnosis and control of the development of hepatic steatosis in dairy cows in the postparturient period. Acta Veterinaria 66(4): 235-243. [ Links ]
Puppione D.L. 1978. Implications of unique features of blood lipid transport in the lactating cow. Journal of Dairy Science 61(5): 651-659. [ Links ]
Recabarren, S.E. 2003. Anestro postparto: posible intermediación de señales metabólicas sobre la secreción de gonadotropinas en vacas lecheras. En: Universidad de Concepción, http://www.chillan.udec.cl/htm; consulta: septiembre 2004. [ Links ]
Rugeles P.C. 2001. Interrelaciones entre nutrición y reproducción en bovinos. Revista Medicina Veterinaria y Zootecnia Córdoba 6(1): 24-30. [ Links ]
Statistical Analysis System (SAS). 1988. User’s Guide: Statistics. version 7. SAS Institute, Inc. Cary, North Carolina, USA. 1028 p.
Strauch, T.A., D.A. Neuendorff, C.G. Brown, M.L. Wade, A.W. Lewis, D.H. Keisler and R.D. Randel. 2003. Effects of lasalocid on circulating concentrations of leptin and insulin-like growth factor-I and reproductive performance of postpartum Brahman cows. Journal of Animal Science 81(6): 1363-1370. [ Links ]
Van Den Top, A.M., M.J. Geelen, T.H. Wensing, G.H. Wentink, Ath. Vant Klooster and A.C. Beynen. 1996. Higher postpartum hepatic triacylglicerol concentration in dairy cows with free rather than restricted access to feed during dry period are associated with lower activities of hepatic glycerolphosphate acyl transferase. Journal of Nutrition 126(1): 76-85. [ Links ]
West, J.W., B.G. Multinix and T.G. Sandifer. 1991. Effects of bovine somatotropin on physiologic responses of lactating Holstein and Jersey cows during hot, humid weather. Journal of Dairy Science 74(3): 840-851. [ Links ]
Working Group on Lipoprotein Measurement. 1995. Recommendations on lipoprotein measurement. National cholesterol education program. U.S. Department of Health and Human Services, NIH Publication No. 95-3944, Bethesda, MD. 94 p. [ Links ]

