










Services on Demand
Journal
Article
 Spanish (pdf)
Spanish (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mailIndicators
-
 Cited by SciELO
Cited by SciELO -
 Access statistics
Access statistics
Related links
-
 Cited by Google
Cited by Google -
 Similars in
SciELO
Similars in
SciELO -
 Similars in Google
Similars in Google
Share

 Permalink
PermalinkRevista Facultad Nacional de Agronomía Medellín
Print version ISSN 0304-2847
Rev. Fac. Nac. Agron. Medellín vol.64 no.2 Medellín July/Dec. 2011
Efecto de Niveles Crecientes de Nitrógeno no Proteico Dietario en Vacas Lactantes sobre las Concentraciones de Metabolitos Nitrogenados en Orina, Sangre y Leche
Effect of Increasing Levels of non Protein Nitrogen in Dairy Cows on Nitrogen Metabolites Concentrations in Urine, Blood and Milk
Rubén Darío Galvis Góez1 ; Héctor Jairo Correa Cardona2; Sorany Milena Barrientos Grajales3 y Yised Muñoz Castaño4
1 Profesor Asociado. Universidad Nacional de Colombia - Sede Medellín - Facultad de Ciencias Agropecuarias. A.A. 1779, Medellín, Colombia. <rdgalvis@unal.edu.co>
2 Profesor Asociado. Universidad Nacional de Colombia - Sede Medellín - Facultad de Ciencias Agropecuarias. A.A. 1779, Medellín, Colombia. <hjcorreac@unal.edu.co>
3 Zootecnista. Estudiante Maestría en Ciencias Agrarias. Universidad Nacional de Colombia - Sede Medellín - Facultad de Ciencias Agropecuarias. A.A. 1779, Medellín, Colombia. <mileshow04@hotmail.com>
4 Zootecnista. Profesional Independiente. Universidad Nacional de Colombia - Sede Medellín - Facultad de Ciencias Agropecuarias. A.A. 1779, Medellín, Colombia. <ymunozc@unal.edu.co>
Recibido: Septiembre 4 de 2009; aceptado: Agosto 12 de 2011.
Resumen. Con el fin de evaluar el efecto de niveles crecientes de nitrógeno no proteico dietario (NNP) en vacas lactantes sobre las concentraciones de metabolitos nitrogenados en orina, sangre y lehe, se evaluaron 15 vacas Holstein que cursaban los primeros 100 días posparto. Estas se asignaron aleatoriamente a tres tratamientos experimentales que consistieron en el suministro de un suplemento alimenticio comercial al que se le adicionó urea de tal manera que se incrementara el consumo de proteína degradable en rumen (PDR) en aproximadamente 0,0 (T0), 10 (T1) y 15% (T2) respecto a los requerimientos. Para las 15 vacas se determinaron las concentraciones de nitrógeno ureico en sangre (NUS), orina (NUO) y leche (NUL) así como las de amonio en sangre (AS), amonio en orina (AO) y la glicemia (GLIC), utilizando kit comerciales acoplados a espectrofotometría. Se realizó un balance de PDR donde se encontraron diferencias significativas entre los tratamientos (P<0,01). El balance para la PDR representó el 2,8; 9,5 y 13,1% de los requerimientos para T0, T1 y T2, respectivamente. Se encontraron correlaciones positivas entre el NUS y el AS, y entre el NUS y el NUL; mientras que la correlación entre el NUO con el AO fue negativa (P<0,1). Las demás correlaciones exploradas no fueron estadísticamente significativas.
Palabras clave: Amonio, metabolismo del nitrógeno, nitrógeno ureico en leche, nutrición animal.
Abstract. In order to test the effect of growing levels of dietary non nitrogen protein in lactating cows on urine, blood and milk nitrogen metabolites concentrations, fifteen cows Holstein in their first one hundred post calving days were used. They were three different experimental treatments assigned at random, which consisted on the supply a commercial feeding supplement without any addition of urea as a source of NNP (T0); the second treatment (T1) consisted on the supply of the same supplement with an addition of urea, so that the supply of PDR was increased 10% approximately to the requirements; finally, in the third treatment, (T2) consisted on the supply of the same supplement which was added with enough urea so that the supply of PDR was increased 15% approximately to the requirements. In these cows were determined concentrations of ureic nitrogen in blood (UNB), urine (UNU), milk (UNM), ammonium in blood (AB) and urine (AU), and glycemia; through spectrophotometric analysis. Finding positive correlations between UNB and AB and UNM, while that correlation between UNU and AU was negative. The other correlations weren't relevant. In addition, it was made a balance of degradable protein in rumen (PDR) where relevant differences were found among the treatments for the tested variables.
Key words: Ammonia, nitrogen metabolism, milk urea nitrogen.
Las pasturas utilizadas en los sistemas de producción de lechería especializada en el trópico alto de Colombia se caracterizan por su alto contenido de proteína cruda (PC) y bajo contenido de carbohidratos no estructurales (CNE) (Gaitán y Pabón, 2002). Esto se debe posiblemente a los altos niveles de fertilización nitrogenada a la que son sometidas (Messman, Weiss y Ericsson, 1981) y que determina, además, el incremento en la fracción soluble de la PC (fracción a) en detrimento de la fracción potencialmente degradable (fracción b) (Rodríguez, 1998). Esta situación incrementa la incorporación de N en la planta en forma de nitrógeno no proteico (NNP). Bajo estas condiciones el exceso de nitrógeno no puede ser utilizado por los microorganismos ruminales, saturando la capacidad de uso del amoniaco (NH3) por parte de estos; así, el exceso de NH3 pasa a la sangre y es transportado hasta el hígado para ser convertido en urea (Moller, 1996). Esto origina un exceso de amonio y de urea en el líquido extracelular (LEC) que se difunde a todos los órganos del animal, afectando la actividad metabólica del hígado, el ambiente uterino y la sobrevivencia embrionaria (Butler, 1998), e incrementando el nitrógeno ureico en la leche (NUL), situación que afecta la calidad de esta. De igual modo, conduce a aumentos en la excreción urinaria de nitrógeno, teniendo efectos indeseables sobre el medio ambiente Alcaráz et al. (2001a). El objetivo de este trabajo fue identificar las alteraciones en algunos indicadores del metabolismo del nitrógeno como resultado de los excesos de nitrógeno no proteico dietario en vacas lactantes.
MATERIALES Y MÉTODOS
Condiciones generales. El trabajo de campo se realizó en el Centro Agropecuario Paysandú de la Universidad Nacional de Colombia, Sede Medellín, ubicado a 2.400 msnm, con una temperatura promedio de 14 ºC y una humedad relativa promedio de 80%.
Se utilizaron 15 vacas Holstein de segunda o más lactancias que cursaban los primeros 100 días posparto. Diariamente se registró la producción de leche y la suplementación alimenticia de cada vaca. Adicionalmente, se emplearon tres vacas secas canuladas en el rumen para la determinación de la degradabilidad de la proteína cruda de la dieta.
Forraje y dieta. Los animales pastoreaban praderas de kikuyo (Pennisetum clandestinum) sometidas al manejo tradicional con fertilización nitrogenada, además, recibieron suplementación alimenticia acorde con la producción de leche y el número de lactancias. Las 15 vacas se asignaron aleatoriamente a tres tratamientos experimentales los cuales consistieron en el suministro de un suplemento alimenticio comercial sin adición de urea como fuente de NNP (T0); el segundo y tercer tratamiento (T1y T2), consistió en el suministro de un suplemento comercial al que se le adicionó urea de tal manera que se incrementara el consumo de PDR respecto a los requerimientos en 10% y 15% respectivamente.
El consumo de forraje se calculó como la diferencia entre la estimación del consumo total de materia seca y el consumo de materia seca del suplemento. Para la estimación del consumo total de materia seca (CMS) se utilizó la ecuación propuesta por el NRC (2001):
CMS (kg/vaca/día) = (((PV^0,75)*0,0968) + (0,372*LCG) - 0,293)*(1 - EXP(-0,192*(SEL + 3,67))
En donde PV es el peso vivo del animal (kg); LCG es la producción de leche corregida al 4% de grasa; SEL son las semanas en lactancia.
Toma de muestras. Al inicio del experimento las vacas fueron muestreadas con la finalidad de establecer las condiciones iniciales como covariables de la investigación. Luego fueron sometidas a las condiciones experimentales durante cuatro semanas consecutivas, al cabo de las cuales se realizó el muestreó final. A cada uno de los animales se le tomaron muestras de sangre de la vena yugular o de la vena coccígea con tubos al vacío. Las muestras fueron centrifugadas a 5.000 rpm durante 10 min luego de lo cual se separó el suero y se envasó en alícuotas de 2,0 mL; también se tomaron muestras de orina y leche en el ordeño de la mañana.
Análisis químicos en orina sangre y leche. En las muestras de suero se cuantificaron las concentraciones de glucosa mediante el kit glucosa oxidasa/peroxidasa (BioSystems®, Barcelona) y en las muestras de orina, sangre y leche las concentraciones de urea mediante el kit urea/Bun-color (BioSystems®, Barcelona) 11537. Para estimar la concentración de amonio en orina y sangre se utilizó el kit Aquamerck® 1.08024.0001 (Darmstadt R.F.A.), un kit cualitativo basado en la formación de un compuesto coloreado de azul de indofenol.
Dado el carácter cualitativo del test original, se procedió a estandarizar la utilización del kit realizando la lectura calorimétrica en un espectrofotómetro Shimadzu® UV-150-02.
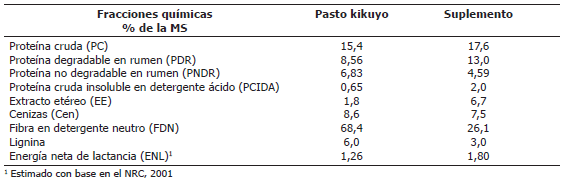
Caracterización nutricional de la dieta. A las muestras de forraje y suplementos alimenticios se les determinó el contenido de cenizas (Cen), extracto etéreo (EE), proteína cruda (PC) y proteína cruda insoluble en detergente ácido (PCIDA). La FDN y la lignina se determinaron por el método de Van Soest (1985) mientras que el contenido de carbohidratos no estructurales se calculó por diferencia (NRC, 2001).
Con los datos de los análisis químicos y degradabilidad, se calculó el contenido de energía neta de lactancia (ENL) del forraje y los suplementos utilizando las ecuaciones reportadas por el NRC (2001).
A las muestras de forraje y suplemento alimenticio se les determinó la degradabilidad ruminal de la materia seca y de la proteína cruda mediante la técnica in situ incubando una bolsa de nylon (de 5,5 x 12,5 cm) con aproximadamente 3,0 g de muestra en cada uno de los tres animales dispuestos para este efecto durante 0, 2, 4, 8, 16, 24, 48 y 72 horas. Una vez cumplidos los tiempos de incubación, las bolsas se enjuagaron con agua de grifo hasta que esta saliera limpia. Las bolsas con muestra se secaron a 60 ºC durante 48 horas, se esperó que su temperatura se estabilizara y luego se pesaron. Para la determinación de la degradabilidad de la materia seca y de la proteína, el residuo resultante de cada bolsa incubada se sometió a la determinación de la materia seca y la PC.
A partir de estos datos se estimaron los parámetros de la cinética de fermentación ruminal para la materia seca (MS) y la PC y la degradabilidad efectiva de estas fracciones (Ørskov y McDonald, 1979). La materia seca y la proteína no degradable en rumen (PNDR) se calcularon como la diferencia entre la MS y la PC total y la MS y la PC degradables en rumen.
Para determinar la degradabilidad efectiva se estimaron las tasas de pasaje del forraje y del suplemento a partir de las ecuaciones propuestas por el NRC (2001), las que se describen a continuación:
Forrajes húmedos: Kp = 3,054+0,614 X1
Donde: Kp = Tasa de pasaje desde el rumen, %/h y X1 = Consumo de materia seca, porcentaje del peso vivo.
Concentrados: Kp = 2,904 + 1,375X1 - 0,020X2
Donde Kp = Tasa de pasaje desde el rumen, %/h, X1 = Consumo de materia seca, porcentaje del peso vivo y X2 consumo de concentrado como porcentaje de la materia seca de la dieta.
La estimación del balance de PDR se calculó por la diferencia entre los requerimientos estimados con el modelo NRC (2001) y el consumo de PDR, calculado a partir del consumo estimado de forraje y el consumo de suplemento y los respectivos análisis de composición química y degradabilidad.
Análisis estadístico. Se realizaron análisis de varianza para establecer las diferencias en cuanto a las concentraciones de urea, amonio y glucosa en las diferentes muestras en que fueron estudiadas (sangre, orina, leche) en función de los tratamientos. Cuando se presentaron diferencias estadísticamente significativas (P<0,05) se adelantó la prueba de Duncan para establecer las diferencias entre medias. Para ello se utilizó el PROC GLM del programa SAS (1998). Asimismo, a través de análisis de correlación se exploraron las relaciones entre el balance para PDR, con las concentraciones de glucosa, urea, y amonio en las diferentes muestras en que fueron analizadas (sangre, orina, y leche). Para esto se eligió el análisis de correlación y se utilizó el PROC CORR del programa estadístico SAS (1998). Para aceptar la significancia estadística de las correlaciones se utilizó el valor de P<0,10, pues se considera que en la exploración inicial de relaciones de asociación, con bajo número de observaciones y cuando no se conocen los coeficientes de variación de las variables implicadas, puede aceptarse este valor.
RESULTADOS
Composición química y contenido energético de los alimentos. En la Tabla 1 se presenta la composición química del forraje y del suplemento experimental.
Tabla 1. Composición química y contenido energético de los alimentos utilizados en estudios del metabolismo en vacas lactantes.
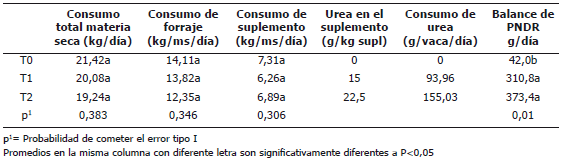
Consumos, requerimientos y balance de PDR. La Tabla 2 muestra los promedios de los consumos de materia seca total, forraje, suplemento y urea para cada tratamiento. No hubo diferencias significativas entre tratamientos para los consumos de materia seca total, forraje y suplemento alimenticio, debido a la alta variabilidad que presentaron los principales factores condicionantes del consumo de materia seca (producción de leche y peso vivo) entre los individuos de cada tratamiento. Los consumos de urea difirieron entre tratamientos, dado que este fue el factor condicionante del experimento. El balance de PDR fue significativamente diferente entre tratamientos.
Tabla 2. Consumos de forraje y suplementos y balance de proteína degradable en rumen en los tratamientos utilizados en estudios del metabolismo en vacas lactantes.
Producción de leche y concentración de metabolitos en sangre orina y leche. En la Tabla 3 se indican los resultados para la producción de leche y las variables metabólicas evaluadas. Se puedo apreciar que ninguna de estas variables fue afectada por los tratamientos.
Tabla 3. Producción de leche y concentración de metabolitos en sangre, orina y leche, en vacas lactantes nutridas con un suplemento alimenticio adicionado con urea.

Correlaciones entre algunas variables metabólicas. En la Tabla 4 se muestran las correlaciones entre algunas de las variables metabólicas entre las que se destacan las correlaciones halladas entre nitrógeno ureico en sangre (NUS) y NUL y entre NUS y GLIC (P<0,05); así como entre nitrógeno ureico en orina (NUO) y amonio en orina (AO); NUL y AS y entre GLIC y NUL (P<0,1). Las demás correlaciones no fueron estadísticamente significativas (P>0,1).
Tabla 4. Correlaciones entre algunas variables consideradas en estudios del metabolismo en vacas lactantes.

Correlaciones entre el balance de PDR y las variables metabólicas. En la Tabla 5 se muestran las correlaciones entre el balance de PDR y las variables metabólicas evaluadas. En esta tabla se aprecia que la concentración de glucosa, la de AS y la de NUO no se correlacionaron con el balance de PDR (P>0,1). Mientras que el NUS, el NUL y el AO se correlacionaron con el balance para PDR (P<0,1).
Tabla 5. Correlaciones entre el balance de PDR y las variables consideradas en vacas lactantes.

DISCUSIÓN
Consumos y balance de PDR. Como se puede observar en la Tabla 2, los consumos de materia seca total, de forraje y de concentrado no presentaron diferencias entre tratamientos, esto fue debido a la distribución aleatoria de los animales, de este modo los principales factores influyentes en la estimación del consumo por el modelo NRC (2001) (peso, producción y días en lactancia) presentaron variaciones similares entre los diferentes tratamientos.
En concordancia con el planteamiento experimental, el balance para la PDR se incrementó en función de la adición de urea en los T1 y T2. Estos balances correspondieron al 2,75, 9,46 y 13,14% de los requerimientos para T0, T1 y T2, respectivamente.
Metabolitos en sangre, orina y leche. Las correlaciones estadísticamente significativas halladas, se presentan entre variables metabólicas relacionadas directamente con el metabolismo del N pero en compartimentos corporales asociados entre sí. De este modo la correlación encontrada entre el NUS y el NUL es explicable por el hecho de que la urea es un metabolito que se difunde fácil y rápidamente entre los diferentes líquidos corporales (Broderick y Clayton, 1997). Montoya et al. (2004) encontraron que la correlación entre el NUS y el NUL en vacas lactantes en un hato del norte de Antioquia fue de 0,65, menor al obtenido en este trabajo. Broderick y Clayton (1997), por su parte, encontraron que la correlación entre estas dos variables fue de 0,91 en tanto que Broderick (1995) la estimó en 0,86, valor similar al logrado aquí.
La correlación entre el AS y el NUL aunque fue estadísticamente significativa, presentó un valor menor al conseguido entre el NUS y NUL. Esto podría ser debido a que las concentraciones sanguíneas de amonio están afectadas por varios mecanismos como la síntesis de urea y la desaminación oxidativa de aminoácidos (Correa y Cuellar, 2004), mientras que la cantidad de urea que difunde de la sangre a la leche se ve afectada principalmente por el gradiente de concentración entre estos dos compartimentos corporales (Madsen et al., 2005).
Normalmente las concentraciones de amonio en la sangre de rumiantes están por debajo de 5 mg/dL (Stanton y Whittier, 2006), de tal manera que cuando los niveles de este metabolito superan dicho valor, se incrementa el riesgo de muerte por intoxicación. Por su parte el Manual Merck (2006) señala que el límite superior a partir del cual pueden comenzarse a encontrar síntomas de intoxicación con amonio es de 2,0 mg/dL. En este trabajo el contenido de este metabolito fue inferior al señalado por ese texto.
La correlación entre el NUO y AO fue negativa (r = -0,398) debido a que en condiciones normales el amonio que pasa del rumen a la sangre es transformado por el hígado en urea, y como tal es eliminado en la orina (Correa y Cuellar, 2004); sin embargo, cuando los niveles de amonio en el rumen se incrementan más allá de la capacidad que tiene el ciclo de la urea de transformar este metabolito en urea, el amonio es transformado en glutamato y, posteriormente, en glutamina, la cual es convertida en α-cetoglutarato en el riñón eliminándose el amonio vía urinaria (Haussinger, 1983; King, 2000). Así, en la medida en que mayor sea la cantidad de amonio que se elimina por la vía del glutamato -; glutamina, menor es su relación con la eliminación por la vía de la urea.
La ausencia de correlación entre las variables relacionadas con el metabolismo del N antes y después de pasar por la filtración renal, podría deberse a que en los nefrones la urea es concentrada para su excreción en la orina. Debido al flujo en contracorriente y a las diferencias en la permeabilidad de las membranas en las asas ascendentes y descendentes de Henle, se crea un gradiente de difusión de la urea en la orina para remover este metabolito desde la sangre (Swenson y Reece, 1993). El flujo de sangre a través de los riñones es constante lo que asegura que la filtración de la urea sea, así mismo, constante independientemente del volumen de orina. Así, con un bajo volumen de orina, la concentración de la urea va a ser mayor que en un alto volumen de orina aunque la cantidad de urea eliminada sea similar (Jonker et al., 1998).
Correlaciones entre el balance de PDR y las variables metabólicas. Aunque la concentración de glucosa en sangre se halla sometida a un mecanismo de control homeostático orquestado principalmente por la insulina, en el caso de los rumiantes este mecanismo es más débil que en los no-rumiantes y, así mismo, es más débil en vacas con altas demandas de glucosa al inicio de la lactancia que la final de la misma (Bauman, 2000). Por ello no es de extrañar la ausencia de correlación con el balance PDR, como ha sido reportado en trabajos anteriores (Galvis et al., 2003).
El NUS y el NUL se correlacionaron con el balance de PDR. Al respecto, Dou et al. (1996) señalan que en la medida en que se incremente el consumo de PC, mayor será la concentración de urea en la sangre. El NUL ha sido utilizado como un indicador de la excesiva fermentación ruminal de la proteína de la dieta (Broderick, 1995). Jonker et al. (1999) revisaron un amplio número de trabajos de investigación en el que se utilizó este metabolito como indicador del estado nutricional de vacas en producción, concluyendo que este metabolito es de alto valor para el estudio del estado proteico de los animales hasta el punto de utilizarse como un predictor del balance proteico (Jonker et al., 1998). En su trabajo Jonker et al. (1998) encontraron que el NUL podría predecir el NUO y, a partir de este último, se podría predecir el nitrógeno ingerido y la eficiencia en el uso del nitrógeno para la síntesis de proteínas lácteas. El NUO, sin embargo, no se correlacionó con el NUS. Según Jonker et al. (1998) la excreción del NUO es de 12,54 g/dL por cada mg/dL de NUL, la pendiente 12,54, representa los 1.254 L de sangre que se estarían filtrando por día a través de los riñones, de los cuales se estaría extrayendo la urea. Este valor en el modelo presentado por Jonker et al. (1998) es, por lo tanto, estimado de manera mecanística a partir de proceso fisiológicos que supuestamente se estarían dando de manera constante en los animales, sin que esto sea realmente así. El modelo parte, así mismo de una premisa incorrecta al asumir que los animales se encuentran en balance cero de N (Alcaraz et al., 2001b). En este sentido, pareciera más probable no encontrar correlaciones entre las variables metabólicas relacionadas con el metabolismo del N y la excreción urinaria del N en cualquiera de sus formas tal y como se discutió anteriormente.
Las correlaciones positivas entre el NUL y el NUS con la PDR podrían explicarse en el hecho de que a mayor PDR, mayor es la cantidad de amonio liberado en el rumen y, por ende, mayor cantidad de urea se sintetiza en el hígado. Así mismo, una mayor cantidad de PNDR en la dieta puede traducirse en mayor concentración de urea en sangre y leche debido a que como lo señala Correa (2006) el balance de aminoácidos de las dietas bajo las condiciones de alimentación que predominan en Colombia, no permiten un uso eficiente de los aminoácidos absorbidos y, por lo tanto, buena parte de los mismos se desaminan en el hígado y terminan formando urea.
El AS no se correlacionó con el balances para PDR debido muy posiblemente a que este es eficientemente transformado en urea o glutamina en el hígado (Correa y Cuellar, 2004). Las correlaciones positivas y significativas entre el AO y los balances proteicos están muy asociadas a la eficiencia en el uso del N consumido, la que bajo las condiciones de alimentación que existen en los hatos lecheros del país, es muy baja. Así, Alcaráz et al. (2001a) hallaron que la eficiencia en el uso del nitrógeno en vacas Holstein lactantes en un hato del oriente antioqueño, fue de tan solo 14,85% en comparación a los valores alcanzados en otras latitudes que son superiores al 20%, consiguiendo promedios del 28% (Jonker et al., 1998). Delgado (2002), Betancur y Trujillo (2004), Saldarriaga y Soto (2004), encontraron valores de eficiencia en el uso del N de 16,9, 16,1 y 19,2%, reafirmando los datos de Alcaraz et al. (2001a) y sugiriendo que buena parte del N consumido se pierde en la orina y, por lo tanto, encontrando altos valores de excreción urinaria en forma tanto de urea como de amonio.
AGRADECIMIENTOS
A la Dirección de Investigaciones de la Universidad Nacional de Colombia, sede Medellín por la financiación de esta investigación (código DIME: 30803874). Al personal operativo del Centro Agropecuario Paysandú por su apoyo y dedicación.
BIBLIOGRAFÍA
Alcaráz, C., D. Alviar y H. Correa. 2001a. Eficiencia en el uso de nitrógeno en vacas lactantes en un hato lechero del oriente antioqueño. Revista Colombiana de Ciencias Pecuarias Suplemento 14: 36. [ Links ]
Alcaráz, C., D. Alviar y H. Correa. 2001b. Evaluación del modelo MUN para la estimación del consumo de materia seca (CMS) en vacas lactantes. Revista Colombiana de Ciencias Pecuarias Suplemento 14: 36. [ Links ]
Bauman, D.E. 2000. Regulation of nutrient partitioning during lactation: homeostasis and homehoresis revisited. pp 311-328. In: Cronje, P.B. (ed.). Ruminant physiology: digestion, metabolism, growth and reproduction. CABI Publishing, New York. 463 p. [ Links ]
Betancur, J.F. y L.G. Trujillo. 2004. Balance de nitrógeno en vacas lecheras de alta producción alimentadas con pasto kikuyo (Pennisetum clandestinum) y dos niveles de suplementación de proteína no degradable en el rumen. Trabajo de grado de Zootecnia, Facultad de Ciencias Agropecuarias. Universidad Nacional de Colombia. Medellín. 30 p. [ Links ]
Broderick, G.A. and M.K. Clayton. 1997. A statistical evaluation of animal and nutritional factors influencing concentrations of milk urea nitrogen. Journal of Dairy Science 80(11): 2964-2971. [ Links ]
Broderick, G.A. 1995. Use of milk urea as an indicator of nitrogen utilization in the lactating dairy cow. In: U.S. Dairy Forage Research Center, http://www.dfrc.wisc.edu/RS95_pdfs/fu5.pdf. 3 p.; consulta: marzo 2009. [ Links ]
Butler, W.R. 1998. Review: Effect of protein nutrition on ovarian and uterine physiology in dairy cattle. Journal of Dairy Science 81(9): 2533-2539. [ Links ]
Correa, H.J., y A.E. Cuellar. 2004. Aspectos claves del ciclo de la urea con relación al metabolismo energético y proteico en vacas lactantes. Revista Colombiana de Ciencias Pecuarias 17(1): 29-38. [ Links ]
Correa, H.J. 2006. Posibles factores nutricionales, alimenticios y metabólicos que limitan el uso del nitrógeno en la síntesis de proteínas lácteas en hatos lecheros de Antioquia. In: Livestock Research for Rural Development. 18 (3): 43. http://www.lrrd.org/lrrd18/3/corr18043.htm; Consulta: noviembre 2008. [ Links ]
Delgado, G.F. 2002. Estudio comparativo del balance de nitrógeno en vacas lactantes de dos grupos genéticos. Trabajo de grado de Zootecnia. Facultad de Ciencias Agropecuarias. Universidad Nacional de Colombia. Medellín. 67 p. [ Links ]
Dou, Z., R. Kohn, J. Ferguson, R. Boston and J. Newbold. 1996. Managing nitrogen on dairy farms: an integrated approach I. Model description. Journal of Dairy Science 79(11): 2071-2080. [ Links ]
Gaitán, S. y J.D. Pabón. Evaluación energética y proteica de los forrajes utilizados en un hato lechero del oriente antioqueño según el NRC 2001. Trabajo de grado de Zootecnia. Facultad de Ciencias Agropecuarias, Universidad Nacional de Colombia, Sede Medellín. 2003. 55 p. [ Links ]
Galvis, R.D., H.J. Correa y N. Ramírez. 2003. Interacciones entre el balance nutricional, los indicadores del metabolismo energético y proteico y las concentraciones plasmáticas de insulina, e IGF-1 en vacas en lactancia temprana. Revista Colombiana de Ciencias Pecuarias 16(3): 237-248. [ Links ]
Haussinger, D. 1983. Hepatocyte heterogeneity in glutamine and ammonia metabolism and the role of an intercellular glutamine cycle during ureogenesis in perfused rat liver. European Journal of Biochemistry 133(2): 269-275. [ Links ]
Jonker, J.S., R.A. Kohn and R.A. Erdman. 1998. Using milk urea nitrogen to predict nitrogen excretion and utilization efficiency in lactating dairy cows. Journal of Dairy Science 81(10): 2681-2692. [ Links ]
Jonker, J.S., R.A. Kohn and R.A. Erdman. 1999. Milk urea nitrogen target concentrations for lactating dairy cows fed according to national research council recommendations. Journal of Dairy Science 82(6): 1261-1273. [ Links ]
King, M.W. 2000. Nitrogen metabolism and the urea cycle, http://web.indstate.edu/themedicalbiochemistrypage.org/nitrogen-metabolism.html. 12 p. Consulta: mayo 2009. [ Links ]
Madsen, T.G., L. Nielsen and M.O. Nielsen. 2005. Mammary nutrient uptake in response to dietary supplementation of rumen protected lysine and methionine in late and early lactating dairy goats. Small Ruminant Research 56(1): 151-;164. [ Links ]
Manual Merck. 2006. Nonprotein nitrogen poisoning: introduction. (ammonia toxicosis). In: The Merck Veterinary Manual, http://www.merckvetmanual.com/mvm/index.jsp?cfile=htm/bc/212400.htm. consulta: noviembre 2008. [ Links ]
Messman, M.A., W.P. Weiss and D.O. Erickson. 1981. Effects of nitrogen fertilization and maturity of bromegrass on nitrogen and amino acids utilization by cows. Journal of Animal Science 70(2): 566-575. [ Links ]
Moller, S. 1996. ''Protein penalty'' in cows consuming pasture. Cattle Practice 4(1): 71-77. [ Links ]
Montoya, N.F., I.D. Pino y H.J. Correa. 2004. Evaluación de la suplementación con papa (Solanum tuberosum) a vacas Holstein lactantes. Revista Colombiana de Ciencias Pecuarias 17(3): 241-249. [ Links ]
National Research Council. 2001. Nutrient requirements of dairy cattle. 7th revised edition. National Academy Press, Washington D.C. 39 p. [ Links ]
Ørskov, E.R. and I. Mc.Donald. 1979. The estimation of protein degradation in the rumen from incubation measurements weighted according to rate of passage. The Journal of Agricultural Science 92: 499-503. [ Links ]
Rodríguez, D. 1999. Caracterización de la respuesta a la fertilización en producción y calidad forrajera en los valles de Chiquinquirá y Simijaca (Estudio de caso). Trabajo de grado, Facultad de Medicina Veterinaria y de Zootecnia, Universidad Nacional de Colombia. Sede Bogotá. 59 p. [ Links ]
Rueda, S., L. Taborda y H.J. Correa. 2006. Relación entre el flujo de proteína microbiana hacia el duodeno y algunos parámetros metabólicos y productivos en vacas lactantes de un hato lechero del oriente Antioqueño. Revista Colombiana de Ciencias Pecuarias 19(1): 27-38. [ Links ]
Saldarriaga, C. y S. Soto. 2004. Efecto de dos edades de rebrote del pasto kikuyo (Pennisetum clandestinum) sobre el balance de nitrógeno en vacas Holstein de alta producción. Trabajo de grado de Zootecnia. Facultad de Ciencias Agropecuarias. Universidad Nacional de Colombia. Medellín. 30 p. [ Links ]
SAS. 1998. STAT Users Guide: Statistics, Version 8. Cary, NC. [ Links ]
Stanton, T.L. and J. Whittier. 2006. Urea and NPN for cattle and sheep. In: Colorado State University Cooperative Extension -; Agriculture, Document, 1.608. http://www.ext.colostate.edu/PUBS/LIVESTK/01608.html. 3 p.; Consulta: noviembre 2008. [ Links ]
Swenson, M.J. and W.O. Reece. 1993. Water balance and excretion, pp. 573-604. In: Duke's Physiology of Domestic Animals. 11th edition. Cornell University Press, New York. 962 p. [ Links ]
Van Soest, P.J. and J.B. Robertson. 1985. Analysis of forrage and fibrous foods. A laboratory manual for animal science. Cornell University Press, New York. 503 p. [ Links ]

