










Services on Demand
Journal
Article
 English (pdf)
English (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mailIndicators
-
 Cited by SciELO
Cited by SciELO -
 Access statistics
Access statistics
Related links
-
 Cited by Google
Cited by Google -
 Similars in
SciELO
Similars in
SciELO -
 Similars in Google
Similars in Google
Share

 Permalink
PermalinkActualidades Biológicas
Print version ISSN 0304-3584
Actu Biol vol.33 no.95 Medellín July/Dec. 2011
RESEARCH PAPERS
EVALUATION OF ETHANOL PRODUCTION FROM TWO RECOMBINANT AND A COMMERCIAL STRAINS OF SACCHAROMYCES CEREVISIAE (FUNGI: ASCOMYCOTA) IN SUGAR-CANE MOLASSES AND REJECTED-BANANA JUICE FROM URABÁ, COLOMBIA
EVALUACIÓN DE LA PRODUCCIÓN DE ETANOL POR DOS CEPAS RECOMBINANTES Y UNA COMERCIAL DE SACCHAROMYCES CEREVISIAE (FUNGI: ASCOMYCOTA) EN MELAZA DE CAÑA DE AZÚCAR Y MOSTOS DE BANANO DE RECHAZO DE URABÁ, COLOMBIA
Carolina Peña-Serna1,2; Carolina Castro-Gil2,4; Carlos A. Peláez-Jaramillo1,5
1 Grupo Interdisciplinario de Estudios Moleculares. Universidad de Antioquia. Medellín (Antioquia), Colombia. cpenaser@gmail.com.
2 Unidad de Biotecnología Vegetal. Corporación para Investigaciones Biológicas. Universidad de Antioquia. Medellín (Antioquia), Colombia.
Correos electrónicos: 4 <carobs23@hotmail.com>; 5 <cpelaez@matematicas.udea.edu.co>.
Recibido: febrero 2011; aceptado: octubre 2011.
Abstract
The production of bioethanol using Saccharomyces cerevisiae (Fungi: Ascomycota) is influenced by sugar concentrations and the fermentation substrate. For that reason, in this study the kinetics of biomass production, residual sugar and ethanol production of four S. cerevisiae strains were evaluated in two fermentation media (sugar-cane molasses and rejected-banana juice) at two sugar concentrations (100 and 170 g/l). The Ethanol Red® and GG570- CIBII strains exhibited the greatest ethanol production, with peak values of 119.74 (35 h) and 62 g/l (15 h), Yps 0.75 and 0.43 g/g, and Qp 3.42 and 2.61 g/l/h, respectively, at 170 g/l of sugar in the sugar-cane molasses broth. In additional, the GG570-CIBII strain showed an increase of 37.1 g/l ethanol with respect to the control strain.
Keywords: adhII gene, bioethanol, pdc gene, recombinant Saccharomyces cerevisiae, rejected-banana juice, sugar-cane molasses.
Resumen
La producción de bioetanol a partir de Saccharomyces cerevisiae (Fungi: Ascomycota) está influenciada por la concentración de azúcares y el sustrato de fermentación. Por ello, en este trabajo se evaluaron las cinéticas de producción de biomasa, azúcares residuales y producción de etanol de cuatro cepas de S. cerevisiae en dos medios de fermentación (melaza de caña de azúcar y banano de rechazo) a dos concentraciones de azúcares (100 y 170 g/l). Las cepas Ethanol Red® y GG570-CIBII presentaron mayor producción de etanol con pico de producción de 119,74 (35 h) y 62 g/l (15 h), Yps 0,75 y 0,43 g/g y Qp 3,42 y 2,61 g/l/h, respectivamente a 170 g/l de azúcares en melaza de caña de azúcar. Adicionalmente, la cepa GG570-CIBII mostró un incremento de 37,1 g/l de etanol con respecto a la cepa control.
Palabras clave: bioetanol, gen pdc, gen adhII, jugo de banano de rechazo, melaza de caña de azúcar, Saccharomyces cerevisiae recombinante.
INTRODUCTION
Bioethanol, used as an oxygenator for gasoline, is produced by the fermentation of sugars present in renewable materials (FNB 2007, Soliclima 2007). Colombia produces around 1.05x106 l/day of bioethanol from sugar-cane juice and molasses (FNB 2007, MME 2007).
This process could increase its efficiency through the use of fermentative microorganisms that produce greater ethanol yields. For that reason, the Plant Biotechnology Unit of the Corporación para Investigaciones Biológicas (CIB) has developed strains of S. cerevisiae that are genetically modified by the insertion of optimized pdc and adhII genes from Zymomonas mobilis, which have shown greater ethanol yields in glucose as a carbon source (Vásquez et al. 2007) than CBS8066 (parental strain).
The enzymes pyruvate decarboxylase (PDC) EC 4.1.1.1 and alcohol dehydrogenase (ADH) EC 1.1.1.1 are present in microorganisms such as S. cerevisiae and Z. mobilis. These enzymes are important for ethanol production (Gunasekaran and Chandra 1999) and have some differences according to the microorganism of origin. For instance, it has been found that the enzymes of Z. mobilis exhibit high affinity for their respective substrates (Brenda database 2007) and that Z. mobilis produces a better ethanol yield than S. cerevisiae (Davis et al. 2006), with values of 0.49 and 0.46 g/g, respectively, in broth cultured with 100 g/l of glucose at 200 rpm, with a pH of 5 and a temperature of 30 ºC.
It is necessary to search for alternative substrates to sugar cane derivatives that may allow actual ethanol production to increase. Taking into account, that Colombia is an exporter country of the banana Cavendish valery of which about 9,877.4 tons/ month are rejected and remain in cultivation fields, leading to contamination (Afanador 2005). Thus, rejected bananas are potential substrate for the production of bioethanol.
In this research, the kinetics of biomass production, residual sugars and ethanol production of two genetically modified yeast strains generated by insertion of optimized Z. mobilis pdc and adhII genes, were evaluated after being cultivated in broths prepared with either sugar-cane molasses or rejected-banana juice.
MATERIALS AND METHODS
Microorganisms and conservation. Fermentations were carried out with four strains of Saccharomyces cerevisiae: parental strain CBS8066 used as a control; recombinant strain GG570-CIBI which harbors the pdc gene of Z. mobilis under the control of the PGK (phosphoglycerate kinase from S. cerevisiae) gene promoter and terminator; recombinant strain GG570-CIBII which harbors the pdc gene of Z. mobilis under the control of the PGK promoter and terminator and the adhII gene of Z. mobilis under the control of the ADHI (alcohol dehydrogenase I from S. cerevisiae) gene promoter and terminator, both strains developed at the Plant Biotechnology Unit of CIB (Vásquez et al. 2007); and a commercial strain, Ethanol Red® (Fermentis).
Yeast strains were stored in a glycerol-YPD culture broth mix at -70 °C in 1.5 ml Eppendorf tubes (Manikandan et al. 2008, Sharma et al. 2007).
Fermentation substrate. Two substrates were used for fermentation, one made from sugar-cane molasses and the other from rejected-banana. Sugar-cane molasses (characteristics shown in table 1), were purchased at the local market from Mayagüez sugar refining industry. Molasses were diluted to the suitable sugar concentration and filtered.
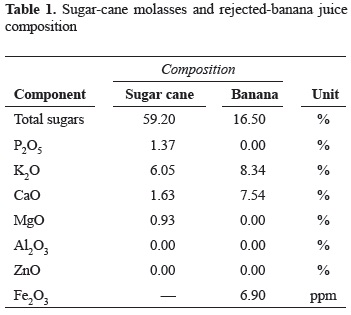
Ripe Cavendish valery banana fruits, rejected from ''La Manzana'' plantation located in Turbo (Antioquia) with composition as shown in table 1, were manually washed, peeled and weighted. Pulp juice was extracted by compression, added hot diluted Ca(OH)2 (100 ml per kg of banana) during gently stirring, allowed for solid agglomeration for 20 min and centrifuged. Banana juice was neutralized with citric acid 0.5% until pH 5.5 and filtered.
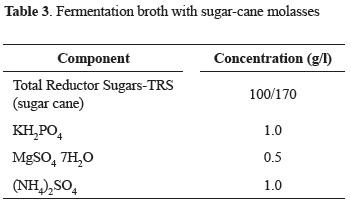
Culture broth. Three different culture broths were used: modified YPD broth for the inoculum preparation and two industrial broths for the fermentation; the fermentation broths were prepared with either, sugar-cane molasses or rejected-banana juice and the pH adjusted to 5.0. The composition of each culture broth is shown in the following tables (2-4):


Inoculum preparation. The inoculum production was prepared in 1000-ml Erlenmeyer flasks with 400 ml of modified YPD broth. 1 ml of conserved strain was added to the sterile broth and incubated at 30 ºC, for 30 h, without mechanical agitation and aeration supplied with an aquarium pump. Once the amount of initial biomass for fermentation was reached (DO660 = 1.3) with viability and vitality above 95%, the supernatant was removed by centrifugation at 4000 rpm.
Fermentation conditions. The fermentations were carried out in 500-ml Erlenmeyer flasks with 400 ml of fermentation broth in orbital shaker at 30 °C, 150 rpm, initial pH of 5 and an inoculum concentration of 8 g/l over 35 h, taking samples of 5 ml every 5 h.
Evaluation of cellular biomass production. Cellular biomass was quantified by the dry weight method (Manikandan et al. 2008, Monsalve et al. 2006, Peña and Arango 2009).
Evaluation of residual sugars. Content of residual sugars was determined using dinitrosalicylic acid (DNS) with previous hydrolysis with 10% HCl at 95 °C (del Rosario and Pamatong 1985, Ergun and Mutlu 2000, Monsalve et al. 2006, Peña and Arango 2009).
Evaluation of ethanol production. Measurement of ethanol production was accomplished by gas chromatography, using an agilent gas chromatograph model 6890, with a flame ionization detector and autosampler (Peña and Arango 2009).
Kinetic parameters determination. Specific growth rate (μx) at exponential growth phase is equal to the maximum growth rate (μmax) (Doran 1995):

Where:
X = cellular biomass concentration at time (t)
X0 = cellular biomass concentration at time (t = 0)
μ = specific cellular growth rate, considered as μmax at the exponential phase
The observed cell yield (Y'x/s) and the observed ethanol yield (Y'p/s) were calculated using curves of cellular growth vs. residual sugar and ethanol production vs. residual sugar, which have slopes of Y'x/s and Y'p/s, respectively (Doran 1995).
The volumetric ethanol productivity, Qp, was calculated using an ethanol production vs. time curve, in which Qp is the slope (Doran 1995).
Statistical analysis. These experiments were developed with a random design, with a balanced factorial arrangement 4 x 22, three repetitions and repeated measurements at 0, 5, 10, 15, 20, 25, 30, and 35 h of fermentation. The response variables were biomass production, residual sugar amount and ethanol production.
RESULTS AND DISCUSSION
Evaluation of biomass production. The fermentation-kinetics experiment was carried out over 35 h, reaching a maximum peak of cellular biomass between 30 and 35 h (figure 1).
In figure 1A, the strains (with the exception of the commercial strain) have a lag phase from 0 to 5 h at the beginning of fermentation and an exponential-growth phase between 5 and 30 h, followed by the stationary phase. Ethanol Red® was the strain that produced the lowest amount of biomass, with a maximum of 12 g/l, whereas recombinant GG570-CIBI produced the greatest biomass, with 15 g/l.
On the other hand, when the strains were cultivated in sugar-cane molasses at 100 g sugar/l (figure 1B), no lag phase neither stationary phase was observed, and the biomass production was slower than the biomass produced in the 170 g/l sugar fermentation process.
During the first 10 h (figure 1C), the different strains, with the exception of the commercial strain, showed an adaptation phase with a decrease in biomass production, which could suggest that the consumption of the substrate was difficult, most likely due to the presence of an inhibitor compound in banana juice. That compound could prevent sugar consumption, causing a decrease in biomass by the autoconsumption of cells. After 10 h, the exponential growth phase began, followed at 30 h by the death phase. In contrast, the commercial strain showed an exponential-growth phase from 0 to 25 h, finally reaching the stationary phase after 35 h.
During cultivation with 100 g sugar/l in banana broth (figure 1D), the strains showed an adaptation phase during the first 5 h with a decrease in the biomass, as was explained previously. Later, the exponential-growth phase lasted until 25 h, when cellular death began.
In general, the strains exhibited greater biomass production in higher sugar concentrations (figures 1A and 1C), due to greater carbon-source availability. In contrast, when the substrates with sugar-cane molasses (figures 1A and 1B) and rejected-banana juice (figures 1C and 1D) were compared, the strains displayed better biomass production when they were cultivated in broth containing sugar cane-molasses than in broth containing rejectedbanana juice (between 11-15 g biomass/l and 8-11 g biomass/l, respectively).
According to statistical analysis, the evaluated factors and their second-order interactions (such as substrate*time, time*sugar and sugar*strain) are statistically significant, with p < 0.0001, whereas the interactions of the third and fourth order, with p > 0.05, do not exhibit a statistically significant effect.
Additionally, statistically significant differences were found between the two substrates (sugarcane molasses and banana juice, with sugar cane molasses proving the better substrate) and between the sugar concentrations (with an initial concentration of 170 g/l generating higher biomass production).
Evaluation of residual sugars. In figure 2 is shown the kinetics of residual sugars during the fermentation. The ANOVA test shows that the factors and their interactions (with the exception of substrate*sugar, sugar*strain, substrate*time*sugar and time*sugar*strain, with p > 0.05) are statistically significant, with p < 0.001.
Statistically significant differences were observed for the factors of substrate type and sugar concentration, while strains did not show statistically significant differences between control and recombinant GG570-CIBI strains and between commercial and recombinant GG570-CIBII strains.
Evaluation of ethanol production. When there is a higher sugar concentration in the fermentation broth, ethanol production is improved (figure 3) because when the fermentation broth contains high sugar concentration (above 3-30 g/l, depending on the strain; Thatipamala et al. 1992), even under high dissolved-oxygen concentrations (Lei et al. 2001), the yeast changes its oxidative metabolism to oxidoreductive or fermentative metabolism, producing a higher ethanol concentration. This phenomenon is known as the Crabtree effect (Converti et al. 1985, Lei et al. 2001, Thatipamala et al. 1992).
When sugar-cane molasses was used (figures 3A and 3B), the strains produced greater ethanol concentration than they did when rejectedbanana juice was used. Furthermore, as noted previously, greater biomass was produced by the sugar-cane molasses substrate. That could suggest that the banana broth contains some kind of inhibitor (Sánchez and Cardona 2008), or, perhaps, the sugars contained in the broth are less available; that is, could be oligosaccharides that cannot be taken up by the yeast, causing a decrease in cellular growth and ethanol production.
Another difference between fermentation broths is that when sugar-cane molasses was used (figures 3A and 3B) the strains produced the maximum ethanol concentration (production peak) in a shorter time (between 15 and 20 h) than with banana broth (figures 3C and 3D), which exhibited production peaks at 30 to 35 h.
According to statistical analysis, ANOVA test suggests that the factors and their interactions are statistically highly significant (p < 0.0001). In addition, the analysis showed statistically significant differences between evaluated factors; that is, the substrate factor that took two values (banana and molasses substrates) presented significant statistical differences, and the sugar-cane molasses substrate demonstrated higher ethanol production than did the rejected-banana juice. Using sugar-cane molasses, the commercial strain (followed by recombinant GG570-CIBII), with 170 g/l of initial sugar concentration produced the best ethanol production.
The recombinant GG570-CIBII showed the best ethanol production comparing to the other recombinant and parental strains, when it was cultivated under high iron concentration (above 150 mM), it means when the strain was grown in sugar-cane molasses at high sugar concentration (figure 3A). Under this condition, recombinant GG570-CIBII increased ethanol production by 37.1 g/l compared to the control (CBS8066) strain.
Peña et al. (2010) showed that the ADHII enzyme encoded by adhII gene of Z. mobilis inserted in the recombinant strain GG570-CIBII, needs the iron ion as an enzymatic cofactor (Gunasekaran and Chandra 1999, Mackenzie et al. 1989), leading to an improvement in ethanol production with respect to the control strain.
On the other hand, Cazetta et al. (2007) reported an ethanol concentration of 2.94 g/l after 24 h of fermentation with sugar-cane molasses broth at 180 rpm and 25 °C, using a Z. mobilis ATCC 29191 strain. Meanwhile, recombinant strains GG570-CIBI and GG570-CIBII produced 41 and 56 g ethanol/l, respectively, after 25 h with the same substrate. This result shows that the recombinant strains are more productive when cultivated in an industrial substrate; moreover, such strains are not inhibited by the presence of salts, which is a problem with Z. mobilis (Gunasekaran and Chandra 1999).
Another study that used an industrial strain of S. cerevisiae in banana broth produced 0.116 l ethanol/ kg fruit after 72 h of fermentation (Hammond et al. 1996), whereas with recombinant GG570- CIBII and commercial strains, 0.035 and 0.04 l ethanol/kg fruit were obtained, respectively, after 35 h. Strains used in the foregoing study reached a higher level of ethanol production than the strains evaluated in this work. However, the production happened in a shorter time period and, additionally, with a lower initial sugar concentration.
Kinetic parameters. The mmax data (table 5) show that the strains increase their specific growth velocity when there is a higher sugar concentration and, additionally, when the strains are cultivated in sugar-cane molasses. These findings are similar to those presented in figure 1.
Regarding to Yxs, the biomass produced per gram of substrate sugar decreases when the strains are grown in sugar-cane molasses broth at high sugar concentration 170 g/l of sugars (except for the control strain) (table 5), which was expected, due to the Crabtree effect.
In contrast, the strains grown in banana broth produced larger amounts of biomass per gram of substrate when the sugar concentration was increased (except for the commercial strain). This result suggests that, with this substrate, the strains used the sugars mostly for cellular growth and not for ethanol production.
Concerning to Yps, the ethanol concentration per gram of sugar increased with the highest sugar concentration evaluated for each strain (except for recombinant GG570-CIBI strain in banana culture and the control strain in molasses broth), meaning that the efficiency of ethanol production was better under high sugar concentration. Moreover, the sugar-cane molasses broth is more propitious for ethanol production than banana broth, and the highest ethanol concentrations were obtained with the commercial strain and the recombinant GG570- CIBII strain (table 5).
Additionally, strains improve ethanol productivity when the fermentation broth contains higher sugar concentration and, in addition, when sugar cane-molasses was used as a carbon source (with the exception of the recombinant strain GG570-CIBII with 100 g sugars/l; table 5).
In general, the strains improve their volumetric productivity by approximately 1 g/(l-h) when the sugar concentration rises from 100 to 170 g/l, with the exception of control and commercial strains in banana broth that showed an increase of 2 g/(l-h) and the recombinant GG570-CIBII strain in sugar-cane molasses broth that showed an improvement of 1.5 g/(l-h).
The highest Qp was observed in sugar-cane molasses medium with 170 g/l of sugar, and the highest productivities were produced by the commercial strain, with a Qp of 3.42 g/l/h, followed by recombinant GG570-CIBII, with a Qp of 2.61 g/l/h.
CONCLUSIONS
Evaluation of kinetics of biomass production, residual sugars and ethanol production showed that the best initial sugar concentration and fermentation substrate are: 170 g/l and sugar-cane molasses compared with 100 g/l and rejectedbanana juice. According to the results, the strains Ethanol Red® and GG570-CIBII exhibited the greatest ethanol production, with peaks of 119.74 (35 h) and 62 g/l (15 h), Yps 0.75 and 0.43 g/g and Qp 3.42 and 2.61 g/l/h, respectively, at 170 g/l of sugar in sugar-cane molasses broth. In addition, the recombinant strain GG570-CIBII showed an increase of 37.1 g/l ethanol with respect to the control strain. This could improve ethanol production at industrial level once the same genetic transformation of GG570-CIBII strain is made in an industrial strain.
ACKNOWLEDGMENTS
This work was supported by a grant from the Agricultural and Rural Development Ministry of Colombia.
REFERENCES
1. Afanador A. 2005. El banano verde de rechazo en la producción de alcohol carburante. Revista EIA, 3: 51-68. [ Links ]
2. Brenda database (Internet). 2007. Brenda - The comprehensive enzyme information system [Internet]. Department of Bioinformatics and Biochemistry TU Braunschweig. Accessed: 16th May 2011. Available from: http://www.brenda-enzymes.org/. [ Links ]
3. Cazetta M, Celligoi M, Buzato J, Scarmino J. 2007. Fermentation of molasses by Zymomonas mobilis: Effects of temperature and sugar concentration on ethanol production. Bioresource Technology, 98: 2824-2828. [ Links ]
4. Converti A, Perego P, Lodi A, Parisi F, Del Borghi M. 1985. A kinetic study of Saccharomyces strains: performance at high sugar concentrations. Biotechnology and Bioengineering, 27: 1108-1114. [ Links ]
5. Davis L, Rogers P, Pearce J, Peiris P. 2006. Evaluation of Zymomonas based ethanol production from a hydrolysed waste starch stream. Biomass and Bioenergy, 30: 809-814. [ Links ]
6. Del Rosario E, Pamatong F. 1985. Continuous-flow fermentation of banana fruit pulp sugar into ethanol by carrageenan-inmobilized yeast. Biotechnology Letters, 7: 819-820. [ Links ]
7. Doran P. 1995. Bioprocess Engineering Principles. London: Academic Press. p. 439. [ Links ]
8. Ergun M, Mutlu SF. 2000. Application of a statistical technique to the production of ethanol from sugar beet molasses by Saccharomyces cerevisiae. Bioresource Technology, 73: 251-255. [ Links ]
9. FNB (Federación Nacional de Biocombustibles) (Internet). 2007. ABC de los alcoholes carburantes. Accessed: 27th May 2011. Available from: http://www.fedebiocombustibles.com/v2/main-pagina-id-5.htm. [ Links ]
10. Govindaswamy S, Vane L. 2007. Kinetics of growth and ethanol production on different carbon substrates using genetically engineered xylose-fermenting yeast. Bioresource Technology, 98: 677-685. [ Links ]
11. Gunasekaran P, Chandra K. 1999. Ethanol fermentation technology-Zymomonas mobilis. Current Science, 77: 56-68. [ Links ]
12. Hammond J, Egg R, Diggins D, Cable C. 1996. Alcohol from bananas. Bioresource Technology, 56: 125-130. [ Links ]
13. Lei F, Rotboll M, Jorgensen SB. 2001. A biochemically structured model for Saccharomyces cerevisiae. Journal of Biotechnology, 88: 205-221. [ Links ]
14. Mackenzie K, Eddy C, Ingram L. 1989. Modulation of alcohol dehydrogenase isoenzyme levels in Zymomonas mobilis by iron and zinc. Journal of Bacteriology, 171: 1063-1067. [ Links ]
15. Manikandan K, Saravanan V, Viruthagiri T. 2008. Kinetic studies on ethanol production from banana peel waste using mutant strain of Saccharomyces cerevisiae. Indian Journal Biotechnology, 7: 83-88. [ Links ]
16. MME (Ministerio de Minas y Energía) [Internet]. 2007. Los biocombustibles en Colombia (Internet). 2007. Ministerio de Minas y Energía. Accessed: 7th August 2011. Available from: http://www.minminas.gov.co/minminas/downloads/UserFiles/File/hidrocarburos/Programa.pdf. [ Links ]
17. Monsalve J, Medina V, Ruíz A. 2006. Producción de etanol a partir de la cáscara de banano y de almidón de yuca. Revista Dyna, 150: 21-27. [ Links ]
18. Peña C, Arango R. 2009. Evaluación de la producción de etanol utilizando cepas recombinantes de Saccharomyces cerevisiae a partir de melaza de caña de azúcar. Revista DYNA, 76: 153-161. [ Links ]
19. Peña C, Arango RE, Restrepo LF. 2010. Efecto de adición de iones hierro y zinc sobre la producción de etanol de dos cepas recombinantes de Saccharomyces cerevisiae. Revista Colombiana de Biotecnología, 12: 158-168. [ Links ]
20. Pleassas S, Bekatorou A, Koutinas A, Soupioni M, Banat I, Marchant R. 2007. Use of Saccharomyces cerevisiae cells immobilized on orange peel as biocatalyst for alcoholic fermentation. Bioresource Technology, 98: 860-865. [ Links ]
21. Sánchez O, Cardona C. 2008. Trends in biotechnological production of fuel ethanol from different feedstocks. Bioresource Technology, 99: 5270-5295. [ Links ]
22. Sharma N, Kalra K, Oberoi H, Bansal S. 2007. Optimization of fermentation parameters for production of ethanol from kinnow waste and banana peels by simultaneous saccharification and fermentation. Indian Journal Microbiology, 47: 310-316. [ Links ]
23. Soliclima. 2007. El etanol como biocombustible (Internet). 2006. Soliclima energía solar. Accessed: 20th May 2011. Available from: http://news.soliclima.com/divulgacion/biomasa/el-etanol-como-biocombustible. [ Links ]
24. Thatipamala R, Rohani S, Hill GA. 1992. Effects of high product and substrate inhibition on the kinetics and biomass and products yields during ethanol batch fermentation. Biotechnology and bioengineering, 40: 289-297. [ Links ]
25. Vásquez J, Castaño H, Marín P, Rodriguez E, Arango R. 2007. Ingeniería genética en rutas metabólicas de Saccharomyces cerevisiae para incrementar la productividad de etanol. Memorias del Sexto Simposio Internacional de alcoholes y levaduras. de congreso. Bogotá (Colombia). [ Links ]
26. Zhu S, Wu Y, Yu Z, Zhang X, Wang C, Yu F, Jin S. 2006. Production of ethanol from microwave-assisted alkali pretreated wheat straw. Process Biochemistry, 41: 869-873. [ Links ]

