











 Spanish (pdf)
Spanish (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mail Cited by SciELO
Cited by SciELO  Cited by Google
Cited by Google  Similars in
SciELO
Similars in
SciELO  Similars in Google
Similars in Google 
 Permalink
PermalinkINTRODUCCIÓN
La diversidad biológica y el estudio de la ecología de las poblaciones han sido un tema central para promover y ejecutar objetos de conservación del entorno natural (MADS y PNUD, 2014). Colombia, como país megadiverso, ocupa los primeros puestos en cuanto a diversidad de especies (Andrade, 2011). En el caso de las mariposas, Colombia posee un total de 3276 especies, dentro de las cuales 350 son endémicas; la riqueza de estos organismos ubica al país como el segundo con mayor diversidad después de Perú (Henao y Gantiva, 2020).
Lepidoptera constituye el tercer orden de insectos con más especies descritas después de Coleoptera y Diptera, contribuyendo a la dinámica ecosistémica al ser partícipes de procesos mutualistas como la polinización o formando parte de la cadena trófica (Villalobos y Salazar, 2020). Además, las mariposas son usadas como bioindicadores del estado de conservación del entorno natural a causa de su sensibilidad a los cambios ambientales (contaminación de fuentes hídricas, cambio climático, extensión de la frontera agropecuaria e implementación de cultivos de uso ilícito) que generan la destrucción del hábitat, los cambios en la cobertura, el uso del suelo y la desaparición o desplazamiento de especies (Andrade, 1998; Morales, 2011) (Sánchez, 2002; Tacuma, 2020), por lo que la abundancia y diversidad de algunas especies de estos insectos deja al descubierto la salud de un entorno debido a que estos factores están directamente relacionados con su estado de conservación.
Adicionalmente, la aceleración de estos procesos antrópicos en regiones apartadas o inexploradas por sus condiciones de difícil acceso o de conflicto, hace prioritaria la necesidad de conocer la diversidad de flora y fauna con el fin de proponer en esos territorios, alternativas de restauración ecológica a mediano y largo plazo. Lo anterior debido al aumento de la deforestación en algunas áreas de Colombia después del posconflicto, condición que puede afectar la conservación de los organismos objeto de esta investigación (Clerici et al., 2020).
Por lo antes expuesto, este estudio tiene por objetivo conocer la diversidad de papilionoideos en la vereda de San Andrés de Pisimbalá, municipio de Inzá, departamento del Cauca y así facilitar la promoción de acciones que contribuyan a su sostenibilidad, teniendo en cuenta el potencial ambiental, turístico y económico que se proyecta en la región. Además, este estudio se convertirá en una herramienta adicional con la que se busca promover el turismo sostenible en la región.
MATERIALES Y MÉTODOS
Área de estudio
El corregimiento de San Andrés de Pisimbalá (02°34´57’’N y 76°02´42’’W) está ubicado en el municipio de Inzá, a una distancia de 110 km de la ciudad de Popayán, sobre la vertiente oriental de la cordillera Central, a una altitud de 1702 m s. n. m. La topografía es variada, resultado de un relieve heterogéneo con una geomorfología de montañas ramificadas casi verticales con crestas agudas y de laderas cortas (Pachón et al., 1996). Sus suelos son húmedos por causa de la presencia de afloramientos hídricos y de acuerdo con Holdridge (1967) es una región con vegetación tropical, por lo que se considera una transición de bosque húmedo a muy húmedo/montano bajo (Bmh-Bh/MB) (Figura 1).

Recolección del material biológico
Se llevaron a cabo seis salidas de campo durante los meses de enero a junio del año 2019, con una duración de cuatro días por muestreo. Se establecieron cuatro transectos de 200 m a lo largo de diferentes hábitats: Transecto 1: Potrero, Transecto 2-4: Cultivos de pancoger y Transecto 3: Bosque secundario, donde se emplearon muestreos con red entomológica y trampas tipo van Someren Rydon (Andrade et al., 2013; Villarreal et al., 2006).
Muestreo con red entomológica
Se realizaron muestreos libres, en un horario de 8:00 a 11:00 y de 15:30 a 18:00 (esfuerzo de muestreo por persona, un total de 6 h de observación y captura por día. Este muestreo lo realizaron tres personas) para completar el registro de especies que presentan hábito crepuscular (Pollard y Yates, 1996). Se procuró siempre que el esfuerzo de muestreo fuera con el mismo horario y personal. Se registró el tiempo muestreado en cada cobertura y la hora en que se recorrió.
Muestreo con trampas tipo Van Someren Rydon
Se instalaron diez trampas dentro de cada una de las coberturas identificadas; como cebo se utilizó fruta fermentada (banano y piña). Se realizaron transectos de 200 m en cada cobertura, donde se instalaron diez trampas separadas entre sí cada 20 m. Las trampas fueron colgadas entre uno y tres metros por encima del suelo, dependiendo de la estructura de la vegetación y altura del dosel. Las trampas se activaron por espacio de 48 h, revisándose cada tres horas (Daily y Ehrlich, 1995) con un esfuerzo de muestreo por transecto de 480 h.
Todos los ejemplares capturados fueron sacrificados por medio de cámara letal, usando acetato de etilo, con el propósito de no deteriorar el individuo. Las mariposas se guardaron en sobres de papel mantequilla con sus respectivos datos (localidad, departamento, municipio, vereda, lugar, fecha de recolecta, hora, nombre del colector, altitud y coordenadas geográficas tomadas con un GPS directamente en campo) para su posterior montaje y análisis (Andrade et al., 2013).
Identificación taxonómica del material
Se utilizaron los patrones de coloración, forma alar, las bandas y algunas estructuras especiales como palpos, patas y órganos genitales como caracteres taxonómicos definidos para la determinación de los ejemplares colectados (Andrade et al., 2013).
En cuanto a la determinación taxonómica, se utilizaron las claves, ilustraciones y descripciones de Andrade (1990, 1995, 2002), Constantino (1995), D’Abrera (1984, 1987a, 1987b, 1994, 1995), García et al. (2002), Le Crom et al. (2002), Le Crom et al. (2004). Además, se siguió la clasificación taxonómica de Lamas (2004) propuesta en Butterflies of America (Warren et al., 2017). La recolección del material biológico se realizó bajo la Resolución 0152 del 12 de febrero de 2015 emitida por la Autoridad Nacional de Licencias Ambientales - ANLA por el cual se otorgó un permiso marco de recolección de especies silvestres de la diversidad biológica con fines de investigación científica no comercial. El material debidamente montado y etiquetado se depositó en la colección entomológica del Museo de Historia Natural de la Universidad del Cauca (MHN-UCC) en la ciudad de Popayán.
Análisis de datos
En primer lugar, se elaboró el listado de las especies encontradas en la zona de estudio; también se realizaron gráficos para representar las familias, géneros y especies más dominantes. Para el análisis de la eficiencia del muestreo, se construyó la curva de interpolación y extrapolación basada en la cobertura de la muestra, que es la proporción del total de individuos en una comunidad representada por las especies observadas (Chao y Jost, 2012). Además, se determinó la diversidad verdadera por medio de los números efectivos de especies (Jost, 2006). Se calculó la diversidad de orden cero (0q) que equivale a la riqueza de especies, orden uno (1q) equivalente a la frecuencia de especies sin tener en cuenta si son comunes o raras (exponencial de Shannon) y orden dos (2q) que representa las especies dominantes (inverso de Simpson) siguiendo el método propuesto por Jost (2006) y Chao y Jost (2015).
Todos los análisis se realizaron por medio del software R Studio versión 1.1.463 (R Core Team, 2021). La diversidad verdadera y la curva de interpolación y extrapolación basada en la cobertura de la muestra se calcularon con los paquetes estadísticos iNEXT (Hsieh et al., 2013) y SpadeR (Chao et al., 2015) y paquetes complementarios como knitr (Xie, 2014) ggplot2 y dplyr (Wickham, 2016). Por último, se calculó el índice de similitud de Bray-Curtis con los paquetes vegan (Oksanen et al., 2020) y stats (R Core Team, 2021).
RESULTADOS
Se registró un total de 128 especies distribuidas en 99 géneros, cinco familias (Nymphalidae, Hesperiidae, Pieridae, Riodinidae y Lycaenidae) y 551 individuos (Anexo 1), representando el 4% de las especies registradas para Colombia por Andrade (2011).
Basado en el método de interpolación/extrapolación, la cobertura de la muestra para los tipos de coberturas vegetal estudiadas arrojó valores que oscilan entre el 48% y 85%, revelando que es posible encontrar más especies de mariposas según aumente el esfuerzo de muestreo (Tabla 1, Figura 2). Entonces, el bosque secundario registra una cobertura de muestra del 85%, seguido de cultivos de pancoger (80%) y potreros (48%).
Tabla 1 Integridad del muestreo
| Tipo de Cobertura | n | S. obs | SC |
|---|---|---|---|
| Bosque secundario | 333 | 106 | 0,85 |
| Cultivos de pancoger | 178 | 67 | 0,8 |
| Potrero | 40 | 29 | 0,48 |
n: abundancia, S. obs: especies observadas, SC: cobertura de la muestra.

La familia Nymphalidae predominó con mayor riqueza y abundancia de especies (54/363) lo que representa un 42% del total registradas, seguida de Hesperiidae (40/77), Pieridae (16/66), Lycaenidae (12/23) y por último Riodinidae (6/22) (Figura 3).

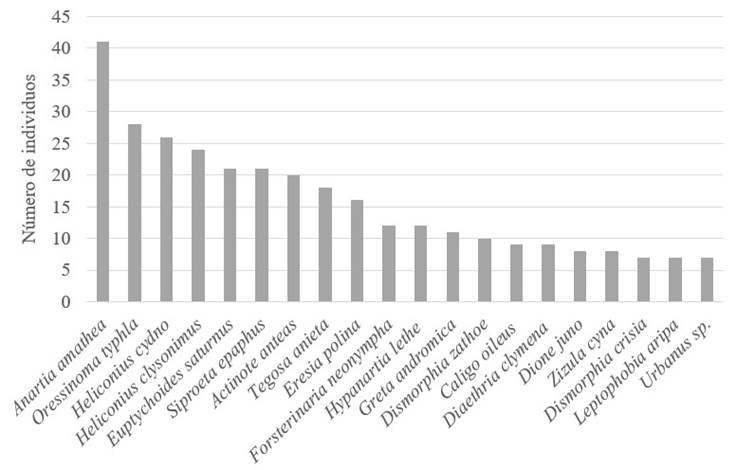
Las especies más dominantes fueron Anartia amathea (Linnaeus, 1758) con 41 individuos, Oressinoma typhla E. Doubleday, [1849] (28), Heliconius cydno (E. Doubleday, 1847) (26), Heliconius clysonymus Latreille, [1817] (24), Euptychoides saturnus (A. Butler, 1867), Siproeta epaphus (Latreille, [1813]) (21) y Actinote anteas (E. Doubleday, [1847]) con 20 registros (Figura 4).
La cobertura vegetal con la mayor representatividad de especies fue el bosque secundario donde se registraron 106 especies y 333 individuos, seguida de los cultivos de pancoger con 67 especies y 178 individuos y potrero con 29 especies y 40 individuos (Figura 5).

Figura 5 Valores relativos del número de especies e individuos de papilionoideos para cada tipo de cobertura.
En cuanto a la diversidad verdadera se encontró que en el bosque secundario el orden 0q (riqueza de especies) fue mayor respecto a las demás coberturas; intermedia en cultivos de pancoger y baja en potrero. De igual manera se registran en ese mismo orden de coberturas un número de especies frecuentes respecto al orden de diversidad 1q. Sin embargo, con la medida de diversidad 2q se encontró que las coberturas de cultivos de pancoger y potreros son similares debido a que presentan un número semejante de especies dominantes (Tabla 2, Figura 6).
Tabla 2 Diversidad verdadera para cada tipo de cobertura
| Tipo de Cobertura | 0 q | 1 q | 2 q |
|---|---|---|---|
| Bosque secundario | 106 | 64,63 | 42,28 |
| Cultivos de pancoger | 67 | 42,17 | 27,27 |
| Potreros | 29 | 26,04 | 22,85 |
0q diversidad de orden 0, 1 q diversidad de orden 1, 2 q diversidad de orden 2.

Figura 6 Diversidad verdadera para cada tipo de cobertura. Las áreas sombreadas señalan los intervalos de confianza del 95% correspondiente a cada tipo de hábitat.
El índice de Bray-Curtis indica que se conformaron tres grupos teniendo en cuenta los tipos de cobertura. El primero está conformado entre bosque y cultivos con una similitud del 52,8%, donde se comparten 36 especies, siendo las más comunes Caligo oileus C. Felder y R. Felder, 1861, Tegosa anieta (Hewitson, 1864), H. clysonimus, O. typhla y S. epaphus. Por último, el grupo hermano adyacente al anterior conjunto es el potrero con una similitud del 16%, en cuando a la estructura de papilionoideos, respecto a las demás coberturas (Figura 7).

DISCUSIÓN
La efectividad de la captura de mariposas para las coberturas de bosque secundario y cultivos de pancoger estuvo bien representada, con más del 80% de la comunidad, siendo el potrero donde se registró el menor número de especies, esto posiblemente a las condiciones climáticas adversas (precipitaciones altas) que afectaron el registro de especies en estas zonas (<48%).
En San Andrés de Pisimbalá no se han realizado estudios sobre biodiversidad de mariposas, por lo que la riqueza registrada durante la investigación representa el primer registro sobre la caracterización de estos organismos. Sin embargo, se han encontrado mayor número de especies en otros estudios realizados sobre la cordillera central como el de Arias y Huertas (2001) con 144 especies y Ospina et al. (2015) con 266 especies registradas.
La mayor riqueza y abundancia de papilionoideos en la zona de estudio estuvo representada por especies de la familia Nymphalidae; esta familia se encuentra ampliamente distribuida en el país tal como lo reportan en sus trabajos Campos y Andrade (2007); Fagua et al. (1999), Orozco et al. (2009) y Prieto y Constantino (1996). Además, Campos y Andrade (2007) enfatizan en el hecho de que dicha familia es muy numerosa en el trópico debido a que se compone de un mayor número de subfamilias, géneros y especies representando el 31% de mariposas del Neotrópico y el 30 % de especies registradas para Colombia. Sumado a esto, la incidencia de una amplia gama de especies generalistas dentro de esta familia permite explorar diversos recursos alimenticios, mostrando así una ventaja adaptativa frente a otras especies pertenecientes a familias como Riodinidae, Lycaenidae y Hesperiidae donde el registro de recursos no es tan amplio (DeVries, 1987; Ospina y Reinoso, 2009).
La especie más abundante fue A. amathea; esta es una especie reportada en toda Colombia, tanto en bosque seco como húmedo hasta los 2000 m s.n.m (Orozco et al., 2009). Esta distribución permite reconocerla como indicadora de áreas abiertas e intervenidas (Tobar et al., 2002; Valencia et al., 2005). Su abundancia se puede atribuir a su tipo de alimentación en estado adulto, ya que es una mariposa que aprovecha el néctar de flores de plantas ornamentales de familias como Acanthaceae, Asteraceae, Melastomataceae, Rubiaceae, Lamiaceae, Solanaceae y Verbenaceae que fueron registradas en la zona (Andrade, 2002; DeVries, 1987; Orozco et al., 2009).
La estructura vegetal heterogénea de la zona de estudio permite el establecimiento de especies comunes de interiores de bosque, borde de bosque y cafetales bajo sombrío como H. cydno, H. clysonimus, Colobura dirce (Linnaeus, 1758), Diaethria clymena (Cramer, 1775), Dione juno (Cramer, 1779), Junonia evarete (Cramer, 1779), O. typhla, Oxeoschistus simplex A. Butler, 1868, Leptophobia aripa (Boisduval, 1836), S. epaphus, T. anieta; cuya presencia se asocia con especies de Passifloraceae, Urticaceae, Amaranthaceae, Annonaceae, y Asteraceae (Carrero et al., 2013; DeVries, 1987; Valencia et al., 2005). Además, estas coberturas presentan un alto grado de humedad al estar asociadas a fuentes hídricas cercanas como la quebrada de San Andrés y la Virgen, donde especies como Rhetus dysonii (Saunders, 1850) y Lieinix nemesis (Latreille, [1813]) liban en las orillas arenosas o cerca de ellas (Le Crom et al., 2004). Adicionalmente, es importante destacar el registro de dos especies indicadoras de áreas conservadas como C. oileus (Palacios et al., 2018) y Episcada polita Weymer, 1899 (Andrade et al., 2007), que se encontraron en zonas de sucesión vegetal secundaria tardía, siendo la cobertura con la más alta conectividad biológica observada.
También es de resaltar que en algunas zonas del bosque secundario se encontraron especies indicadoras de áreas abiertas como A. amathea, A. anteas, S. epaphus, E. polina, Altinote stratonice (Latreille, [1813]), Castilia eranites (Hewitson, 1857) y D. juno (Andrade, 2002; Andrade et al., 2007; Carrero et al., 2013; Palacios et al., 2018; Salazar et al., 2010; Tobar et al., 2002). Lo anterior puede deberse a que esta cobertura presenta, en algunos sectores, un mayor nivel de luminosidad por la entresaca selectiva (Prieto y Constantino, 1996), generando cambios en el microclima y las condiciones físicas tales como régimen de luz, temperatura y humedad (Ospina, 2014). Circunstancias que favorecen la colonización de plantas de la familia Asteraceae que es característica de hábitats abiertos, intervenidos y zonas de transición (Casas et al., 2017; Vélez et al., 2015) y frecuentada por mariposas nectarívoras. Algunos caminos de herradura al interior de esta cobertura presentaban materia orgánica en descomposición producto de los desechos de animales como vacas y caballos, al igual que cebos naturales como frutos caídos lo que generaba el registro de un amplia variedad de mariposas generalistas (Prieto y Constantino, 1996) como Zizula cyna (W. H. Edwards, 1881) que adicionalmente visita flores de Lantana camara (Andrade et al., 2007; Salazar et al., 2010) observada en el sitio o Pronophila unifasciata Lathy, 1906 que es considerada generalista y no requiere condiciones ambientales específicas (Coral y Pérez, 2017; Orozco et al., 2009).
En cuanto a los perfiles de diversidad, es importante mencionar que el bosque secundario y los cultivos de pancoger presentan una mayor riqueza de especies efectivas en comparación con el potrero que arrojó el valor más bajo; probablemente, esto se debe a que existe una mayor oferta alimenticia en el bosque secundario y los cultivos de pancoger tanto para especies generalistas como especialistas. Esto contrasta con otros trabajos donde se registra la mayor diversidad en este tipo de coberturas (Coral y Pérez, 2017; Millán et al., 2009). Respecto al orden 1q se destaca a C. oileus, T. anieta, H. clysonimus, O. typhla y S. epaphus quienes son las especies más frecuentes en bosque y cultivos. Esto posiblemente se deba a las condiciones medio ambientales de estas coberturas que favorecen su tránsito y reproducción (Ríos, 2007). En términos de dominancia (orden 2q), en las tres coberturas muestreadas se registraron especies dominantes como A. amathea y O. typhla. De igual manera Rueda y Andrade (2016) encontraron que algunas áreas abiertas como cultivos y potreros tienen una dominancia de mariposas relativamente altas, posiblemente por la amplia abundancia de brinzales y latizales que ofrecen recursos como plantas hospederas y alimenticias.
En la zona de estudio es evidente la diferencia de riqueza y abundancia de papilionoideos entre las coberturas vegetales, debido a que estos parámetros dependen de las condiciones físico-biológicas del hábitat que determinan la tolerancia fisiológica de las mismas, generando dinámicas de depredación y competencia (Simonson et al., 2001). No obstante, se observa una similitud del 52,8 % entre bosque secundario y cultivos de pancoger en cuanto a la estructura de mariposas como consecuencia de la interacción entre hábitats adyacentes, ya que ambas coberturas se componen de vegetación heterogénea y representan una gran parte de la zona de estudio. Sumado a esto, la actividad agrícola que se destaca en la zona es la producción de café bajo sombrío, por lo que se pueden hallar cafetales mezclados con otro tipo de vegetación y aledaños a relictos o fragmentos de bosque, que según Coral y Pérez (2017) favorecen una mayor presencia de especies de lepidópteros, generando así una mayor conectividad biológica en la región.
Finalmente, se concluye que las condiciones del área de estudio, a pesar del desarrollo de actividades antrópicas como expansión de zonas agrícolas y pastoriles, aún permiten sustentar una amplia diversidad de especies de papilionoideos que deben ser objeto de estudio y conservación por parte de la comunidad. Se debe destacar que el reconocimiento de la diversidad de estas especies en una zona de interés cultural y ecoturística para el país contribuirá con información de línea base para apoyar propuestas encaminadas hacia un turismo sostenible que se viene dando en la región. También se resalta la importancia del presente estudio como uno de los pioneros sobre el conocimiento de la biodiversidad en Tierradentro, vereda de San Andrés de Pisimbalá.



