











 English (pdf)
English (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mail Cited by SciELO
Cited by SciELO  Cited by Google
Cited by Google  Similars in
SciELO
Similars in
SciELO  Similars in Google
Similars in Google 
 Permalink
PermalinkIntroduction
The physiography of the Colombian Caribbean is composed of geomorphic units associated with high coasts, such as hills and massifs, elevated abrasion platforms, marine terraces, eroded rock formations, and glacis or inclined planes, as well as units associated with coastal lowlands including coastal plains, floodplains, estuaries, coastal lagoons, mudflats, dunes, beaches, pins, rods, bars, and reef platforms (Ingeominas, 1996). Depending on rain annual amount and distribution, there are 14 climate units in three provinces: arid to dry in the north, dry to humid in the central area, and humid to very humid in the south. The Tayrona National Natural Park, Santa Marta, department of Magdalena, Colombia (between 11°18' and 11°21' N and 74°4' and 74°7' W) is a low massif with ecogeographic components such as estuaries, beaches and cliffs, lake environments, and hills (El Cielo at 500 m of altitude).
Geological formations in the study area comprise a representation of Tertiary material including Neogene sediments, Miocene and Pliocene rocks, Eocene plutonic and sedimentary rocks, and Paleocene-Eocene volcanic materials. Mesozoic materials are associated with metamorphic rocks from the Upper Cretaceous and Jurassic. The Quaternary sediments are recognizable in flat areas with valleys of colluvial and alluvial influence partly found in the maritime zone forming sedimentary lagoons behind the marine bars. Additional information on the Caribbean region of Colombia and the Tayrona National Natural Park study area can be accessed at www.colombiadiversidadbiotica.com.
The soils have moderately coarse textures (loamy-sandy) on igneous or mixed materials and medium textures (loamy and loamy-silty) on metamorphic and mixed materials (Cortés, 1975). The humidity gradient varies from west to east: very dry (500 mm) near Taganga - Santa Marta, and humid (1400 mm) near Arrecifes - Cañaveral with a transition spot in the Bay of Cinto (Figure 1). The precipitation regime is bimodal and tetra-seasonal with two rainy periods (May to June and September to November) and two dry periods (December to April and July to August). The average annual temperature is 27 °C (Rangel & Carvajal, 2012).

Materials and methods
Study area
Neguanje Bay is located at the central-western coastal zone of the park bordered on the West and East by two South-North oriented hill ranges reaching up to 150 meters which separate it from the Gayraca and Cinto Bays, respectively (Figure 1). The southern limit is defined by high and low hills that gradually descend to the sea and are part of the maritime drainage system of the park hydrographic complex. These hills harbor small ravines and channels that reduced streams and remain dry for most of the year (Carbon-D. & García-Q., 2010).
Cortés-C. (2017) characterized the Caribbean mangrove vegetation and determined the phytosociological arrangement including the Lagunculario racemosae-Avicennion germinantis alliance present in the Neguanje area. The arboreal stratum of these forests is formed by L. racemosa and Hilairanthus germinans but Rhizophora mangle is very poorly represented while the fern Acrostichum aureum grows densely on the herbaceous stratum. The L. racemosae-A. germinantis association also includes other species from the surrounding wet and dry forests, such as Libidibia coriaria, Pithecellobium dulce, Ficus matiziana, and Maclura tinctoria.
The beaches and flooded areas harbor larger thickets communities dominated by Solanum campechiense growing together with Heliotropium indicum, Alternanthera paronychioides, Glinus radiatus, Portulaca oleracea, Mimosa dormiens, Euphorbia hirta, and Tarenaya (Cleome) spinosa. The slopes are covered by forests composed of Handroanthus billbergii, Libidibia punctata, and Capparis (sensu lato) species, as well as communities dominated by Astronium graveolens, which is quite similar in its floristic composition to the forests located in the hills around Santa Marta (Carbonó & García, 2010; Rangel, 2012).
Landscape transformation events have occurred in the swamp including local ones: subsidence and uplift of blocks, ancient coral bars, and coastal terraces (Robertson & Martínez, 1999), as well as global ones: earthquakes and tsunamis in the Caribbean (Quintero, et al., 2020; Bolívar, et al., 2019), and fluctuations in the sea level, regressions and transgressions (Beltrán, 2015), which have influenced on the evolution of the territory. The effects of the tectonics, the variations in the sea surface temperature, and the precipitation have also been reflected in the changes in the stratigraphic and biostratigraphic sequences (Jaramillo, et al, 2012).
Fieldwork and laboratory analysis
In Neguanje (Playa Pozo sector at the Tayrona N.N.P.), we recovered a 4 m-deep column of sediment. We described the core stratigraphy in the field and then sealed and transported the sample in 50 x 5 cm plastic tubes to the palynology and paleoecology laboratory at the Instituto de Ciencias Naturales, Universidad Nacional de Colombia. A portion of the sediments was used for detailed stratigraphic and LOI (loss on ignition) analyses and carbon-14 dating.
We used the acetolysis methodology for the physicochemical preparation to extract the palynomorphs in the sample from the column and the two samples from the surface for recent pollen sedimentation analysis (Erdtman, 1986). The final part of the samples was subjected to flotation treatment with zinc chloride (ZnCl2) (D'Antoni, 2008).
The slides samples for the palynological analysis were prepared using gelatin-glycerin according to the Kisser formula (Brown, 1960). Each sample was studied under a Zeiss Primo Star light microscope and we counted the maximum number of palynomorphs (grains, fungal spores, and ferns) and other palynomorphs (algae, plant, and insect remains). We made the taxonomic identification of the pollen grains based on the atlases of Roubik & Moreno (1991), Jiménez, et al. (2008), and García-M., et al. (2011) and comparing with the reference collections of the Palinoteca del Instituto de Ciencias Naturales, Universidad National de Colombia. In some cases, the microscopic analysis effort included several sheets of one depth to reach a minimum number of palynomorphs (Table 1), which explains the lack of uniformity in the class intervals (depth).
A set of samples were selected and sent to the BETA Analytic Laboratory (Florida) and to the University of Colorado (USA) for carbon-14 dating. The dates (14C ages) supplied by the laboratories followed standardized protocols and were based on the IntCal20 calibration (Reimer, et al., 2020); they appear in the column marked with * in the palynological diagrams. Sedimentation rates were estimated according to the zonation of the palynological diagram. In all cases, both the depth at which the original sample was taken and the width of the intervals of each of the zones appear in the column (Table 2) We conducted sedimentological (microstratigraphic) and main components analyses for the bottom sediments (Jaramillo, et al., 2012, Coronel, 2013).

To determine the reservoir capacity of the sediments, we used the loss of mass by calcination method (LOI) (Dean, 1974). We estimated LOI at 105°C (moisture %), LOI at 550°C (organic carbon %), and LOI at 950°C (inorganic carbon %) to establish the system's organic reservoir capacity also by means of the LOI method (Dean, 1974). According to Eriksoon & Olsson (2015)), the organic matter of continental origin comes from the vegetation near the study site. It may indicate high primary productivity related (in our case) to the extension or reduction of forest vegetation (flooded areas and mainland slopes). Therefore, the marine-origin organic matter is associated with mangrove vegetation; LOI values at 950 °C refer to the carbonate content in the sediments originating from elements of marine origin (corals, shells).

For the palynological diagrams, we used the Tilia software (version 1.7.16; Grimm, 2002) and the elements included in the pollen sum were those from reliable ecological and phytogeographic information sources while the spores of ferns, fungi, and other palyno-morphs whose autecology and other ecological characteristics were not well documented were grouped under the name "elements not included in the pollen sum." Finally, two surface sediment samples (recent pollen rain) were also analyzed.
For the phytoecological segregation of the palynomorphs (grains, spores) found in the sediments, we followed the recommendations of Rangel (2012), Cortés-C. & Rangel-Ch. (2013), and Cortés-C. (2017). We conducted surface sample or current pollen rainfall analyses throughout the entire column inside the palynological diagrams including the elements explained in the next section.
Elements included in the pollen sum
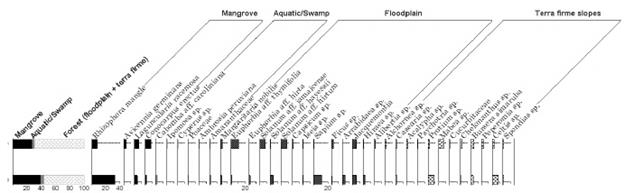
The following elements (genera, species), whose grains appear in the group included in the sum, were selected because their ecological characteristics and geographical distribution patterns reinforce the segregation.
Mangrove vegetation. R. mangle, H. germinans, L. racemosa, Conocarpus erectus, and A. aureum.
Aquatic and swamp vegetation. Cyperus sp., Poaceae, Ambrosia peruviana, and Althernanthera aff. paronychioides. Other elements with values below 1% clustered in the curve were tagged as diverse aquatic-swamp vegetation (Cabomba aff. caroliniana, Ipomoea sp., Scleria sp., Eclipta sp., Persicaria sp. (Polygonum) sp., Amaranthaceae, Hibiscus sp., and Convolvulaceae).
Floodplain vegetation included Mascagnia macradena, Margaritaria nobilis, Phyllanthus sp., Cissus aff. biformifolia, Euphorbia aff. thymifolia, Solanum sp., S. aff. jamaicense, S. aff. umbellatum, Ficus sp., Bactris sp., Jacquemontia sp., and Sapium sp. Other elements were gathered in diverse-floodable areas where their pollen representation was less than 1%; these taxa were represented by Cissus aff. fuliginea, Tournefortia sp., Euphorbia aff. hirta, Solanum aff. hayessii, Capsicum sp., Arrabidaea sp., Iseia sp., Hiraea sp., Alibertia sp.
Slope vegetation of the mainland (tropical dry forest) was composed of Celtis sp., Acalypha sp., Protium sp., Mabea sp., Anacardium sp., Handroanthus sp., Fabaceae, Asteraceae, Prestoea sp., and Arecaceae. Other species were assembled in a curve under the name mainland-diverse slopes. The pollen representation of this group was below 1% and the taxa identified were Spermacoce sp., Sabicea sp., Psychotria sp., Rubiaceae (aff. Psychotria), Alchornea sp., Casearia sp., Croton sp., Sida sp., Faramea sp., Faramea occidentalis, Maripa sp., Geophila sp., Justicia sp., Spondias sp., Schefflera sp., Tabebuia sp., Bursera sp., Cedrela sp., Jatropha sp., Chelonanthus sp., Peperomia sp., Euphorbiaceae, Rubiaceae, Moraceae, Mimosaceae, Bignoniaceae, Boraginaceae, and Bombacoideae (Malvaceae).
Elements not included in the pollen sum
These were pollen grains: Asteraceae (indeterminate) type Jacquemontia, Podocarpus sp.; algae: Pseudochizaea; fern spores: Polypodium (Polypodiaceae); fungi spores: fungi spores, ascospores, and other palynomorphs: phytoclasts, zooclasts, and fungiclasts.
Here we document the major paleoenvironmental changes that occurred in the study area during the last 2000 years based on stratigraphic, sedimentological, and palynological analyses on a 4-m-deep borehole. Our research questions addressed the variations in the floristic composition of the vegetation types and their extension, especially the mangrove swamp, and their relationships with climatic characteristics (precipitation).
Results
Stratigraphy
In the 400-cm-deep core located in Playa Pozo (11°19'-17.02" N; 74°04'- 35.99" W), the stratigraphic sequence corresponded mainly to clays with enriched levels of silt, sandy silt, and sands towards the middle of the column (Figure 2).

Moisture-carbon-loss on ignition (LOI)
We differentiated four areas in the three figures that were used for the description.
Humidity (LOI at 105° C). In zone I, between 400 and 300 cm deep, the values fluctuated between 14% and 30% and up to 40%; in zone II, from 300 to 200 cm, variations were between 5% and 30% with a maximum value of 48%; in zone III, from 200 to 100 cm, values were between 10% and 20% and as high as 40% this being the zone with the lowest humidity in the entire core; in zone IV, between 100 and 1 cm, values ranged from 30% to 40% and up to a maximum of 54%, this being the zone with the highest humidity of the entire nucleus (Figure 3A).
Organic carbon (LOI at 550 ° C). In zone I, we registered values between 2% and 4% and as high as 36%, this being the zone with the lowest values; in zone II (300-200 cm), values ranged from 4% to 10% and reached as high as 24%; in zone III (200-100 cm), values were between 10% and 20% with up to 40%-54%, this being the highest value in this component; in zone IV (100 - 1 cm), values ranged from 4% to 6% and the extreme values reached 20% (Figure 3B).

Figure 3 Moisture and organic and inorganic carbon values (LOI at 105°C, 550°C, and 950°C) from Neguanje core
Inorganic carbon (LOI at 950 ° C). In zone I, values were between 2% and 15% with extreme points of 30% and up to 48%; in zone II, we registered values from 10% to 14% and as high as 40% and 60%; zone III had the highest values ranging between 12% and 16% and as high as 32% and 40% while zone IV had the lowest values, between 4% and 10% (Figure 3C).
Main components and class (origin) of organic matter in sediment
Figure 4A shows the contents (%) of mineral and organic components throughout the sediment column. In the four differentiated zones (the entire nucleus), there is a high content of mineral fraction (between 60% and 80%) with the highest values in zone II.

Figure 4 Main components of the Neguanje core - Playa Pozo. A. Main components = organic matter; mineral fraction. B. Continental organic components. C. Marine organic components
The following was the distribution of organic matter components and their origin (continental or marine) (Figures 4B, 4C):
In zone I, 401 to 302-cm deep, marine origin components were predominant (corals, shells, jaws, fish bones, and an array of calcareous structures), with an average of 21% and extreme variations between 6% and a maximum of 46%, while the components of continental origin (peat, autochthonous and allochthonous coal, roots, stems, and wood) had an average value of 6.3% and values as low as 3%.
In zone II, 301 to 202 cm, there was a slight predominance of continental components with a mean of 9% and extremes of 20%, and an average of 7% with extremes of 12% in marine components.
Zone III, 201 to 102 cm, exhibited a shared domain with continental-origin components of 11.7% on average and maximum values of 23% and marine components with an average of 11.2% and maximum values of 31%.
Finally, in zone IV, 101 to 1cm, continental components predominated with an average of 16% and maximum values of 80% while the marine components registered an average of 6% with extreme values of 14%.
Sedimentation rates
In the deepest part (zone I), the sedimentation rate was slightly higher than that from the superficial zones (IV-V). The highest value was registered in zone II followed by zone III, which was 2.3 and 3.5 times higher than that of the deepest zone (Table 1). These values were also part of the variation gradient (0.06 and 0.66 cm/year) in other mangrove environments such as Laguna Camarones in the department of La Guajira (Castaño, 2004), Cispatá in the department of Córdoba (Palacios, et al., 2012), and La Caimanera in the department of Sucre (Beltrán, 2015).
Recent pollen sedimentation
In the two samples analyzed, we identified 36 palynomorphs belonging to 22 families and 30 genera. Pollen representation was dominated by floodplain vegetation (38%), followed by mangroves (33%), and mainland forest (25%). The swamp vegetation (aquatic-swamp) was represented but in a very low proportion (4%).
As found in groups previously classified, floodplain vegetation included E. aff. thymifolia with an average pollen frequency of 6%, and S. aff. jamaicense and S. aff. hirtum with a value of 4%. Rhizophora mangle dominated in the mangrove swamp (20%) followed by L. racemosa (6%), C. erectus (5%), and H. germinans (2%). In the mainland forest, we found harbor taxa such as Protium (6%), Celtis (5%), and Mabea (3%), and in the swamp vegetation (aquatic-swamp) C. aff. caroliniana, A. peruviana, Ipomoea spp., Cyperus, and Poaceae, with less than 1% of the pollen frequency each (Figure 5).
Palynological diagram
In table 2 we present the palynomorphs (pollen grains, spores, and others) we counted. At some points, not enough materials were recovered and, therefore, the palynological diagrams show discontinuities as the intervals between samples were not constant. Between the surface and the 95-cm depth, the concentration of palynomorphs was low so it was necessary to extend the sampling interval while in most of the other sections of the column sampling was every 5 cm. There were sections in which palynomorphs were not recovered and in other their counts were very low. For example, between 380 and 400 cm, we found only four to seven grains, and, therefore, this material was not used in the analysis.
Zoning, description (palynological diagrams) and reconstruction of the vegetation changes.
Five zones were differentiated. From zones I to IV in the palynological spectrum there is a domain of mangrove-associated elements. In zone V, the outermost (recent) representation of the dry forests of the nearby slopes predominates (Figures 6A, B and 7). A detailed description offers us the following characteristics:
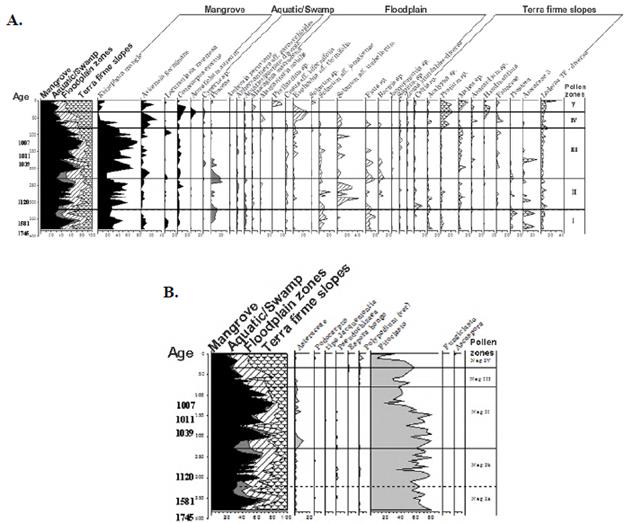
Figure 6. A. Diagram of the elements included in the sum of pollen. B. Elements not included in the pollen sum

Zone I. Depth: 380 - 322cm. Mangrove representation prevailed, especially R. mangle (45%), H. germinans (10%), L. racemosa (5%), and C. erectus (5%) followed by the vegetation of flooded areas with S. aff. jamaicense (20%) and Ficus sp. (20%) and vegetation on nearby slopes (mainland) represented by species of Acalypha (10%), Protium (5%), Mabea (10%), Prestoea sp. (10%), and Arecaceae (20%). The aquatic-swamp vegetation was poorly represented with Poaceae elements (10%). Among those elements not included, phytoclasts reached an 80% representation and the algae genus Pseudochizaea sp., 5%.
Paleoenvironment: Estimated age: 1605-1230 years B.P. The mangrove swamp communities (direct marine influence) were dominated by Rhizophora mangle. In the floodplain (less marine influence), the aquatic and swamp vegetation reached an important representation, especially towards the end of the area. The lower-scrub forests dominated by Solanum and Ficus species were also established, and in the nearby slopes, a mixed palm community dominated with species of the Prestoea, Acalypha, and Protium genera, which increased their distribution area towards the end of the zone. Humidity (precipitation) conditions in the lagoon surroundings were higher compared to those in the present time. Organic components, both marine and those related to the flooding process reached their maximum expression.
Zone II. Depth: 322 - 230 cm. Mangrove representation dominated with R. mangle reaching the highest values (60%) followed by H. germinans (10%), C. erectus (10%), and L. racemosa (5%). The flooded vegetation areas increased compared to zone I with S. aff. jamaicense (20%), Ficus sp. (20%), Bactris sp. (10%), E. aff. thymifolia (5%), and S. aff. umbellatum reaching the maximum value of 40%. Hillside vegetation (firm land) decreased its representation with Mabea sp. (5%), Protium sp. (5%), and three species of Arecaceae (10%). Aquatic-swamp vegetation was poorly represented with Poaceae (20%). The predominance of phytoclasts persisted among the elements not included.
Paleoenvironment: Estimated age: 1230-1064 years B.P. The distribution areas of the mangrove with R. mangle and the scrub-low forest of S. aff. jamaicense, S. aff. umbellatum and Ficus sp. vegetation in the floodplain increased while on the slopes, the area of forest dominated by Acalypha, Protium, and Mabea species decreased. Compared to the previous zone, humidity conditions (precipitation) were slightly lower and the marine organic components fewer.
Zone III. Depth: 230 - 80cm. The predominance of mangrove areas was represented by R. mangle reaching the highest values (80%) of the entire column followed by H. germinans (15%), C. erectus (5%), and L. racemosa (5%). The representation of flooded areas vegetation decreased with the presence of E. aff. thymifolia (10%), S. aff. umbellatum (40%), S. aff. jamaicense (10%), Ficus sp. (20%), Bactris sp. (10%), Mabea sp. (15%), and three Arecaceae species (20%). Aquatic vegetation in swamp areas decreased and they were dominated by Poaceae (10%) and Cyperus sp. (5%). Among the elements not included, phytoclast representation began to decrease towards the end of the zone.
Paleoenvironment: Estimated age: 1064 to 655 years B.P. Mangrove dominance and greatest extension were represented by R. Mangle, H. germinans, and C. erectus. The aquatic-swamp vegetation extension decreased significantly; both the scrub-low forest dominated by Solanum and Ficus species and the forests located on the slopes with Mabea maintained their distribution area. Humidity conditions (precipitation) in the lagoon surroundings were lower than those of the previous zone but the continental and marine organic components increased and their maximum points corresponded to the greatest coverage of the mangrove.
Zone IV. Depth: 80 - 33 cm. Although mangrove areas represented by R. mangle were predominant, they began to decrease drastically towards the top (10%), however, H. germinans (30%) and L. racemosa (10%) increased. Representation of hillside vegetation (mainland) with species of Protium (15%) and Mabea (10%) increased considerably, as well as the flooded-area vegetation with E. aff. thymifolia (20%) and S. aff. umbellatum (10%). Aquatic-swamp vegetation was very poorly represented with Cyperus sp. and Poaceae elements (<5%). Among the elements not included, the predominance of phytoclasts prevailed, although their representation was lower compared to the previous sub-zone.
Paleoenvironment: Estimated age: 655 to 270 years B.P. The area occupied by the mangrove reduced, especially due to the strong decline of R. mangle while the representation of H. germinans and C. erectus increased, mainly towards the end when A. aureum appeared. The areas previously covered by the mangroves were occupied by scrub vegetation with M. macradena, E. aff. thymifolia, M. nobilis, and by floodplain vegetation. Firm land forests on the slope areas were dominated by Protium and Anacardium species extending to the flat areas adjacent to the mangrove. Organic matter of marine origin decreased (mangrove reduction) towards the end of the zone while that of continental origin increased associated with the extension of the flooded vegetation areas. Humidity conditions (precipitation) were lower than those of the previous zone.
Zone V. Depth: 33 - 0 cm. Representation of hillside vegetation (mainland) predominated with Protium sp. reaching up to 20%, Mabea sp. (20%), and other elements such as Handroanthus sp. (10%) and Anacardium sp. (5%). The representation of flooded vegetation areas with E. aff. thymifolia (20%), M. macradena (5%), and Phyllanthus sp. (20%) prevailed. In the mangrove community, R. mangle showed minimal values (15%) compared to those recorded in previous areas; the representation of H. germinans (25%) and C. erectus (20%) increased, and A. aureum (10%) appeared. The low representation of the aquatic and swamp vegetation persisted. Among the elements not included in the pollen sum, again phytoclasts dominated, although to a lesser extent compared to the initial zone (60%).
Paleoenvironment: Estimated age: 270 years B.P. to present time. The area occupied by the mangrove drastically reduced due to the significant regression of R. mangle and H. germinans while C. erectus and A. aureum representation persisted. Hillside forests with Protium, Mabea, and Handroanthus increased their range and reached the basin. Current precipitation values (600-700 mm) qualify the region as very dry (Rangel & Carvajal, 2012). The organic matter of continental origin reached the highest values.
Organic matter components and main palynological diagram relationships
When comparing the LOI organic matter components curves and the main palynological diagram between 550 and 950°C it is possible to differentiate the following situations (Figure 8):

Figure 8 Summary of vegetation changes, main components, continental organic matter, and marine organic matter. Mang: Mangrove; Ac/P: Aquatic-swamp; Inun: Flood plain; TF: Mainland
Zone I: Depth: 401 - 302cm. This includes zone I and the lower part of zone II in the palynological diagram. LOI values at 550 °C were between 2% and 4%, the lowest in the entire column. Marine components predominated, especially shells and other calcareous elements, which relate to LOI values at 950 °C with extreme points of 30% and up to 48%. Mangrove was the vegetation type with the greatest coverage (from 380 cm deep).
Zone II. Depth: 301 - 202 cm. This comprises three-quarters of zone II and the lower part of zone III in the palynological diagram. LOI contents were between 550 and 950° C, similar to the previous zone; organic matter decreased, especially marine components. The vegetation was dominated by mangroves communities.
Zone III. Depth: 201 - 102cm. This comprises three-quarters of zone III in the main palynological diagram. LOI values were at 550 °C (4% to 10%) and 950 °C (4% to 10%), the highest in the entire column. Compared to the previous zone, continental organic matter increased, as well as that of marine origin with an increase of calcareous remains towards the end of the zone. Regarding vegetation, here we found the mangrove area with the greatest coverage and development.
Zone IV. Depth: 101 - 1cm. This includes the upper part of zone III and zones IV and V in the palynological diagram. Compared to the previous zone, the LOI values between 550°C and 950°C decreased as seen in the reduction of marine origin organic matter. In centimeters 1 and 2, the values in continental components (differentiable plant remains) reached an extreme value of 76%. Regarding vegetation, the mangrove area reduced and the area the flooded forest increased.
Discussion
Palynological associations in sediments and affinities with current vegetation types
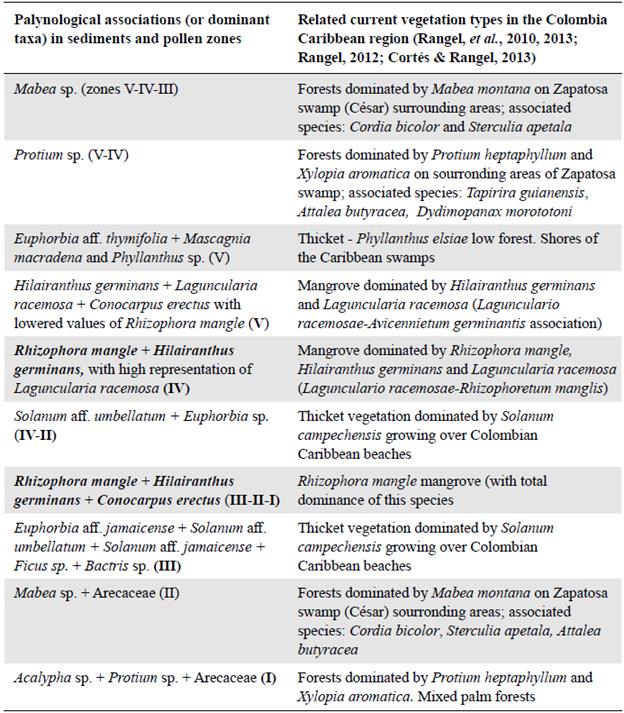
Table 3 shows the relationships between the palynological associations defined in this study based on their respective palynomorphs in the sediment and their correlation with current vegetation types according to their floristic composition. The correspondence grade between mangrove types dominated by R. mangle and by H. germinans was very significant as both types were associated with the same salinity conditions in the water column while the dry forest was dominated by Mabea montana and Protium heptaphyllum.
Palaeoenvironmental changes and their relationship with other Colombian Caribbean areas
For the period from 2000 to 500 years BP, Jaramillo, et al. (2012) and Vélez, et al. (2014) mentioned the presence of the vigorous development of mangrove communities in several localities of the Ciénaga Grande de Santa Marta (Boca de López, Hospitalito, La Ahuyama, Caño Perdiz, Bocas de Aracataca), and Castaño (2004) in Laguna Camarones, Guajira. These communities were dominated mainly by Rhizophora and significant values of Hilairanthus (Avicennia) were also measured. These associations were related to the dry conditions (low precipitation values) predominating in this period in the swamp sector and its surrounding areas.
Table 3 Relationship between the palynological associations detected in the sediment analysis and current vegetation types (Neguanje)
In the Neguanje region, the largest mangrove development (R. Mangle) reached values up to 60% in the pollen spectrum occurring between 1078 and 654 years B.P. (see zone III of the palynological diagram). In the Ostional swamps, Cispatá Bay, this happened ca. 1380 years B.P. (Palacios, et al., 2012), and 838 years A.P. in La Caimanera (Beltrán, 2015).
In the Ciénaga Grande de Santa Marta (C.G.S.M.), sediments were associated with a warming period between 1930 and 1660 years B.P. In Neguanje, the predominance of organic components of marine origin can also be associated with warming conditions. Between 1660 and 900 years B.P., in the C.G.S.M., a shorter warm or "cold" period was detected, which differs from Neguanje, as the presence of organic matter in its sediments of marine components predominated, and we identified a greater dominance and coverage of mangrove vegetation. In the C.G.S.M., Jaramillo, et al. (2012) classified as warm the period between 900 and 570 years B.P. According to Van der Hammen & Troncoso (1992) around 750 years B.P., dry conditions prevailed in the several localities of the lower Magdalena River. In Neguanje, at the end of this period, an increase in organic components of marine origin was observed, which would indicate warm conditions. Between 500 years A.P. and the present, warm or "cold" periods were detected in the CGSM. In this region, a high number of mineral components were found and, in some cases, shell deposition as well. In Neguanje, we identified high contents of the mineral component, a condition that can be present in cold conditions.
Based on the temperature calibration curves of the last 2000 years (D Arrigo, et al., 2006; Christiansen, et al., 2012), and the precipitation and extension data, we adapted the interpretation of the types of vegetation to the Neguanje region as follows:
Zone I. Depth: 380 - 322 cm. Estimated age: 1605-1230 A.P. Compared to current climate variations (27.3°C temperature and 650 mm annual precipitation), the temperature and humidity conditions (precipitation) in the surroundings were slightly higher. Mangrove communities were established in swamp (marsh) areas highly dominated by R. mangle.
Zone II. Depth: 322 - 230 cm. Estimated age: 1230 to 1064 years A.P. Compared to zone I, the temperature was slightly lower and the humidity and precipitation conditions were slightly higher. The mangrove distribution area increased in the swamp (marsh) with R. mangle as the dominating species.
Zone III. Depth: 230 - 80 cm. Estimated age: 1064 to 655 years A.P. This was the zone with the warmest conditions according to the sequences set up in the sediment column. The humidity conditions (precipitation) in the areas surrounding the lagoon were lower than in zone II. We detected the effects of two climatic warming peaks: one towards the base of the area, ca. 1060 years B.P., with an approximate increase of 0.3°C above the global average estimated for the territory (27.3°C). The other peak was registered towards the top part, ca. 940 yr. B.P., with an increase of 0.5°C above the average related to the medieval climate optimum (records of maximum solar activity). In this zone, we recorded the largest extension of R. mangle mangrove. This period can be directly associated with a very strong marine influence as corroborated by an increase in transgression velocity rates.
Zone IV. Depth: 80 - 33 cm. Estimated age: 655 to 270 years B.P. The temperature decreased with respect to zone III. A marked decrease in the water levels was also observed in the swamp due to a lower entry of seawater, a decrease in precipitation around the swamp, and a slight drop in local temperature. It is possible that the decrease in marine influence is associated with a light regression process in the territory. The extension of the mangrove community was considerably reduced, and the exhumation of the territories facilitated the extension of the floodplain scrublands and the forests on the slopes (mainland) occupying areas contiguous to the mangrove.
Zone V. Depth: 0 - 33 cm. Estimated age: 270 years A.P. to the present. The upper part reflected the current conditions of temperature (27.3°C) and precipitation in the territory (650 mm annual precipitation). Compared to zone IV, the temperature was higher (1 to 1.2°C) and precipitation was less. In the swamp (marsh), the extension of the mangrove communities decreased drastically while the area covered by tropical dry forests on the slopes increased.

