










Services on Demand
Journal
Article
 Spanish (pdf)
Spanish (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mailIndicators
-
 Cited by SciELO
Cited by SciELO -
 Access statistics
Access statistics
Related links
-
 Cited by Google
Cited by Google -
 Similars in
SciELO
Similars in
SciELO -
 Similars in Google
Similars in Google
Share

 Permalink
PermalinkUniversitas Psychologica
Print version ISSN 1657-9267
Univ. Psychol. vol.5 no.2 Bogotá Jan./May 2006
ALEJANDRO CASTILLO MORENO* Y ANGÉLICA PATERNINA MARÍN
UNIVERSIDAD DEL NORTE, BARRANQUILLA
Correo electrónico: mcastillo@uninorte.edu.co.
ABSTRACT
In this paper we checked the principal researches and theories to explain the attention system functioning. We are going to start reviewing along time about the concept of attention, from filter theories and resources distributor theories, to the current theories in which attention is conceived as a control system. From this last point of view, we will emphasize on the attentional networks theory of Posner, that proposes different systems to explain diverse aspects of attention, but they are related to each other. At last in this paper, we will mention experimental results that have been important to characterize the selective attentional mechanisms of the human visual system, using the attentional spotlight model for this aim.
Keywords: Attention, attentional system, attentional networks, attentional spotlight, psychopedagogy.
RESUMEN
En este artículo revisamos las investigaciones y teorías que más han contribuido en la explicación del funcionamiento del sistema atencional. Comenzaremos haciendo un repaso histórico del concepto de atención, desde las ideas de filtro y de distribuidor de recursos, hasta las más actuales que la conciben como un mecanismo de control. Desde esta última perspectiva destacaremos la teoría de las redes atencionales de Posner, en la que se proponen sistemas diferentes que explican distintos aspectos de la atención, pero que están íntimamente relacionados. En la última parte del artículo presentaremos aquellos resultados experimentales que han permitido caracterizar al mecanismo atencional selectivo del sistema visual, utilizando para ello la metáfora del foco atencional.
Palabras clave: Atención, sistema atencional, redes atencionales, foco atencional, psicopedagogía.
En la vida cotidiana los humanos recibimos estímulos procedentes de un gran número de objetos y eventos, presentados todos ellos de forma simultánea. Sin embargo, sólo una pequeña parte son relevantes para nosotros. Debemos atender únicamente a aquellos que utilizaremos para alcanzar nuestros objetivos o metas, e ignorar los que pueden ser causa de distracción o interferencia. Los estímulos a los que atendemos los llamamos objetivos y a los ignorados distractores. La atención selectiva es el proceso mediante el cual realizamos esta diferenciación.
Las referencias a procesos atencionales se remontan a la antigüedad (véase Tudela, 1992), pero su estudio sistemático no se produce hasta finales del siglo XIX, cuando W. James (1890) define la atención y establece una taxonomía. Es bien conocido que James enfatiza la función selectiva de la atención (“Es la toma de posesión por parte de la mente, de uno entre los muchos simultáneamente posibles objetos o series de pensamientos.”), aunque también la vincula con la consciencia y los objetivos del individuo. No es de extrañar, por tanto, que el aspecto selectivo de la atención sea una de sus características más estudiadas en la psicología moderna.
En los años 50 y 60 la atención era normalmente conceptualizada como un proceso facilitatorio. Algún mecanismo cerebral seleccionaba los estímulos que debían ser atendidos y la atención facilitaba de alguna manera el procesamiento posterior de tal información. Estas ideas llevaron a plantear teorías basadas en canales de capacidad limitada (Broadbent, 1958; Deutsch & Deutsch, 1963; Treisman, 1964), donde la información pasaba a través de una serie de etapas que solo podían procesar un número limitado de objeto estimulares. Este “cuello de botella” era un filtro donde la información que pasaba era procesada a niveles profundos mientras que el resto simplemente se disipaba en los niveles anteriores. Este tipo de teorías dio lugar a que la experimentación se centrara en cuestiones tales como el lugar donde estaba situado el filtro atencional. Para algunos autores (Broadbent, 1958) (Véase Figura 1) estaba situado en las primeras etapas del procesamiento. Todos los estímulos que llegan al sistema sensorial son analizados según sus características físicas, pero sólo uno atraviesa el filtro cada vez, es seleccionado, y pasa al sistema perceptual de capacidad limitada. La contrastación empírica vino de estudios en los que se intentaba poner de manifiesto deficiencias atencionales sobrecargando el sistema mediante la manipulación de la cantidad de información que se presenta a los sujetos (Cherry, 1953; Posner, 1978) y de experimentos en los que se quería demostrar la serialidad en la identificación de estímulos (Sperling, 1960).

Sin embargo, otros autores consideraron que el filtro se encuentra en estadios de procesamiento posteriores (Deutsch & Deutsch, 1963; Van der Heijden, 1981) (Véase Figura 2). Apoyados en algunas evidencias experimentales provenientes de estudios que utilizaban tareas de escucha dicótica (Lewis, 1970) y tareas Stroop, propusieron que el filtro atencional no actuaba hasta que todos los estímulos (o al menos los más relevantes o familiares de la escena) fueran procesados y categorizados semánticamente. La idea sería que todos los estímulos que llegan a los sistemas sensoriales pasan por todas las etapas de procesamiento hasta alcanzar su representación en memoria (Norman, 1968). Una vez estas han sido activadas (lo que significa que han sido procesadas semánticamente y categorizadas) procesos de arriba-abajo activan más, o seleccionan, la representación mental del estímulo objetivo, con lo que es éste el que pasa a tomar el control de la acción.

Los modelos de filtro enfatizan un aspecto de la atención, su función selectiva, pero tienden a ignorar que esa función es necesaria sólo si se asume que el sistema de procesamiento es de capacidad limitada. Cuando el énfasis se traslada a la capacidad, se produce tanto un cambio en los procedimientos experimentales como en los modelos explicativos. Kahneman (1973) (Véase Figura 3) propuso que los sistemas de procesamiento tienen una cantidad limitada de recursos, los cuales pueden ser usados para la realización de diferentes procesos o tareas. En este modelo, la atención es la encargada de distribuir los recursos entre ellos, para que se ejecuten de manera adecuada, dependiendo del objetivo final del sujeto. Kahneman sugirió el tradicional concepto de arousal (activación general) como un sistema que provee activación o recursos no solo para actividades de carácter físico, sino también para las psicológicas.

En años posteriores comenzaron a surgir teorías que postulaban un segundo proceso que ayudaría al facilitatorio en la tarea de selección. La idea consistía en que, al mismo tiempo que se producía la facilitación, los estímulos distractores presentes en la escena con capacidad para interferir en el normal procesamiento de los objetivos son inhibidos, es decir, su activación es reducida de forma activa. Tanto estudios experimentales como razones teóricas avalaban un mecanismo de este tipo. Entre estas últimas podemos destacar principalmente dos (Houghton & Tipper, 1994). Por un lado se puede argüir la ganancia en velocidad en el proceso de seleccionar los objetivos de entre todos los estímulos que llegan al sistema perceptual. Si, al mismo tiempo que el estímulo objetivo correspondiente está recibiendo activación, los distractores presentes en la escena están siendo inhibidos, o sea, se está reduciendo su nivel de activación, la distancia -en términos activacionales- que los separa aumentará mucho más rápidamente, con lo que la discriminación entre uno y otros se alcanzará antes.
La segunda razón teórica que se puede destacar es la dificultad que en algunos casos podría darse en la selección, si actuara sólo el mecanismo facilitatorio. El nivel de activación de un nodo o neurona debe forzosamente tener dos límites físicos, uno superior y otro inferior. Pues bien, si, por ejemplo, se presentan dos estímulos, uno como objetivo y otro como distractor, que producen una alta activación en sus representaciones o nodos (por ser de brillo intenso, de grandes dimensiones, etc.) una buena discriminación no podría darse con solo activar más a uno de ellos, ya que alcanzaría el nivel máximo de activación rápidamente, sin haber conseguido aún una distancia suficiente para la selección. Sin embargo, con un mecanismo encargado de inhibir el estímulo distractor, esta dificultad no existiría.
Los estudios experimentales que más han aportado en la demostración de la existencia de un mecanismo inhibitorio han utilizado preferentemente tareas tipo Stroop (Dalrymple-Alford & Budayr, 1966; Neill, 1977) y de priming negativo (Castillo & Catena, 2003; Marí-Beffa & Nievas, 2002). Por ejemplo, Neill (1977) presentaba a los sujetos palabras coloreadas y les pedía que respondieran al color de la tinta e ignoraran el significado de la palabra. La presentación se hacía de forma serial. Los resultados mostraron que cuando el sujeto debía responder a un color que era el mismo que designaba la palabra del ensayo anterior, la respuesta era más lenta que cuando no había ningún tipo de relación entre los dos ensayos. Este efecto parecía indicar que existía algún tipo de inhibición del significado de las palabras, lo que les hacía estar menos disponibles si se les necesitaba en una ocasión posterior. Esta demora en el tiempo de reacción ante un objetivo relacionado con un distractor precedente fue denominado priming negativo (Tipper, 1985).
Este tipo de aportaciones llevaron a una reconceptualización de la atención, que pasó a ser considerada como un proceso complejo y flexible, que se adapta a las necesidades y exigencias que plantea al sujeto la tarea que está realizando. Se pasó de la metáfora de “filtro” a otras como “fuente de recursos” (Norman & Bobrow, 1975), “foco” (Posner, Snyder & Davidson, 1980) o “foco con zoom” (Erikssen & Yeh, 1985). La idea de foco atencional se fundamentaba en la existencia de un mecanismo que facilitaba aquellos estímulos objetivos situados dentro de una región específica del mapa perceptual y permitía ignorar los estímulos distractores situados fuera de esta región. La mayor activación de los objetivos les permitía pasar a fases de procesamiento posteriores. Las propiedades y funcionamiento de este supuesto foco atencional han sido aspectos muy estudiados dentro del campo de la atención en los últimos años. Se ha investigado la forma del foco (Eriksen & Eriksen, 1974), el tamaño (LaBerge, 1983; Eriksen & Murphy, 1987), si puede ser dividido (Bichot, Cave & Pashler, 1999, Cepeda, Cave & Bichot, 1998; Kramer & Hahn, 1995; Hahn & Kramer, 1998; Shaw & Shaw, 1977; Sperling & Melchner, 1978; Umiltà, 1995), y algunas otras cuestiones que veremos más adelante.
A continuación vamos a exponer el modelo propuesto por Posner, actualmente considerado como el más sólido y el que más apoyo experimental ha recibido.
Las redes atencionales
En los últimos años la atención ha pasado de ser considerada un mecanismo de procesamiento de la información, a ser considerada un mecanismo central de control de los sistemas de procesamiento (Posner & Dehaene, 1994; Tudela, 1992). Este mecanismo, distribuido en diferentes lugares del sistema nervioso, ejerce sus funciones a través de procesos facilitatorios e inhibitorios. Actualmente se han identificado tres sistemas atencionales separados anatómica y funcionalmente (Posner & Raichle, 1994; Fan, McCandliss, Sommer, Raz, & Posner, 2002; Callejas, Lupiañez & Tudela, 2004): anterior, posterior y sistema de vigilancia. El estudio de estas tres redes nos permitirá revisar lo que actualmente se conoce acerca de los sistemas atencionales del cerebro humano.
El sistema atencional posterior parece estar relacionado con todo lo referente al control del procesamiento espacial (Posner & Raichle, 1994). Sería el encargado de centrar la atención en la posición del campo visual donde está situado el estímulo objetivo. Para ello realiza las operaciones de desenganche de la atención del objeto en el que estaba centrada, movimiento por el campo visual hasta la nueva posición y enganche de la atención en el estímulo designado como objetivo actual. Estas tres acciones son realizadas por mecanismos que están situados en distintas áreas cerebrales, a saber: lóbulo parietal posterior, colículo superior y ciertas áreas talámicas (principalmente el núcleo pulvinar).
Una vez la atención ha cambiado a la nueva localización y el contenido visual de esta zona ha sido transmitido, el sistema atencional anterior entra en juego. La principal función de este mecanismo parece ser la detección de objetos y el reconocimiento de su identidad. Se ha identificado esta segunda red de la teoría de Posner con la llamada Working memory (Baddeley, 1986), y esta, a su vez, se ha asociado con la consciencia. Así, parece que el sistema atencional anterior sería un mecanismo ejecutivo, que haría conscientes el objeto atendido y sus propiedades, y velaría por el cumplimiento de las metas establecidas1 . También este mecanismo parece estar formado anatómicamente por varias áreas cerebrales, principalmente el cíngulo anterior, los ganglios de la base y el córtex dorsolateral prefrontal, es decir, las áreas que forman parte del circuito del ejecutivo central (Funahashi, 2001) (Véase Figura 4).
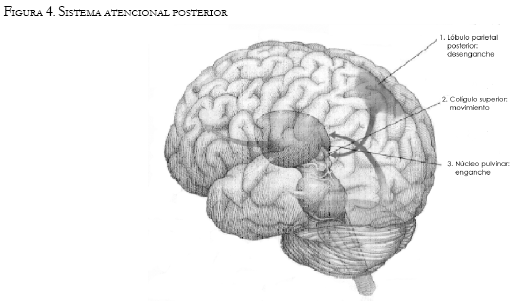
El tercer sistema que forma el mecanismo atencional es el llamado sistema de vigilancia: entra en funcionamiento cuando el sujeto debe estar alerta ante la aparición de un posible estímulo objetivo. Durante este periodo parecen producirse una serie de cambios funcionales en el sistema atencional que preparan el cerebro para la detección rápida del posible objetivo (entre estos cambios cabe destacar la inhibición que sufre el sistema atencional anterior). Anatómicamente el mecanismo atencional de vigilancia está formado por neuronas de norepinefrina del locus coeruleus que proyectan principalmente en áreas de los lóbulos frontal y parietal derechos.
Sistema Atencional Posterior
Este sistema es el encargado de orientar la atención hacia la localización visual donde están situados los posibles estímulos objetivo. La finalidad última de este alineamiento es el aumento en la eficacia del posterior procesamiento de los objetos situados en la posición visual elegida.
La red atencional posterior también parece estar relacionada con la orientación de los sistemas atencionales en memoria, más en concreto con la exploración de imágenes recuperadas de la memoria (Posner, 1980).
Para el estudio de este tipo de orientación de los sistemas atencionales se ha utilizado una tarea que posibilita separar este proceso de otros que están involucrados en operaciones de tipo visual (Véase Figura 5). En un procedimiento típico, un ensayo comienza con un punto de fijación en el centro de la pantalla, donde el sujeto experimental debe centrar su mirada. En las instrucciones se le pide que no mueva de ese lugar sus ojos durante todo el experimento. A continuación aparece un estímulo que indica en qué lugar se presentará con más probabilidad el estímulo objetivo. Esta señal puede ser central o periférica. Una señal central es aquella que indica la posición del objetivo mediante un código determinado (por ejemplo, una flecha apuntando hacia el lugar elegido), mientras que la periférica señala el lugar mediante la posición en la que se encuentra (p.e. un cuadrado blanco en la posición donde se espera aparezca el objetivo). Esta señal se espera que anime al sujeto a mover su atención hacia el lugar indicado con el fin de mejorar el tiempo de reacción en la respuesta. Después de un intervalo de tiempo aparece el estímulo objetivo en una de dos posibles posiciones y el sujeto debe presionar una tecla tan pronto como lo detecte. La posición real del objetivo con respecto a la señalada previamente determina las dos principales condiciones experimentales. El objetivo puede aparecer en el lugar indicado por la señal previa (ensayos válidos) o puede aparecer en el lugar opuesto (ensayos inválidos). Si realmente ha habido, por parte del sujeto, una anticipación en la orientación de la atención hacia donde indica la señal previa, se espera que la respuesta en los ensayos válidos sea más rápida -efecto facilitatorio- mientras que en los ensayos inválidos se espera un enlentecimiento en la respuesta -efecto inhibitorio-. Para medir estos dos efectos, se comparan los tiempos de reacción en estos tipos de ensayos con otros en los que no aparece ninguna señal anticipando la posición del objetivo (ensayos neutrales). En un experimento bajo estas condiciones suelen encontrarse diferencias positivas entre ensayos neutrales y válidos, beneficios, y diferencias negativas entre ensayos neutrales e inválidos, costes. Estos dos efectos aparecen tanto en los tiempos de reacción (Posner, 1980) como en los errores de respuesta (Bashinski & Bachrach, 1980). A causa de esto la tarea ha sido denominada de costos y beneficios.

Posner (Posner & Raichle, 1994) explicó los resultados de esta tarea partiendo de un sistema que realiza tres operaciones cuando hay que mover la atención de una localización a otra: desenganche, movimiento y enganche del foco atencional.
Operación de desenganche. Antes de iniciar el movimiento hacia la nueva localización, la atención debe desengancharse de su antiguo objetivo. Parece que el sistema cerebral encargado de llevar a cabo esta operación está en el lóbulo parietal. Una fuente de evidencia importante son los estudios con pacientes con daño cerebral y estudios que utilizan la Tomografía de Emisión de Positrones (PET). A continuación veremos con detalle estas dos líneas de investigación.
Una lesión en humanos del lóbulo parietal origina el llamado síndrome clínico del neglect, consistente en una incapacidad por parte del paciente de informar de estímulos situados en la parte contralateral a la lesión (DeRenzi, 1982). Este fenómeno también puede surgir cuando hay lesiones en otras partes del cerebro, como en el tálamo, cerebro medio y algunos lugares de la corteza, pero parece que es en el lóbulo parietal derecho donde las lesiones desembocan con más frecuencia en neglect. Cuando estos pacientes realizan la tarea atencional expuesta anteriormente, surgen una serie de resultados que sugieren que el problema se encuentra en la operación de desenganche. Mientras que en los ensayos válidos los tiempos de reacción son similares a los encontrados en sujetos sanos, cuando el paciente con neglect debe responder en un ensayo inválido en el que el objetivo está en el lado contralateral a la lesión, sus tiempos de reacción suelen ser dos o tres veces superiores, e incluso no llegar a responder. Este resultado junto con el hecho de que no existen diferencias en los tiempos cuando la señal es periférica o es central, parece indicar que el problema está en que el sujeto no puede desenganchar su atención una vez la ha fijado en un objeto situado en la misma cara de la lesión. Al igual que en los estudios generales sobre neglect, se ha encontrado que este efecto de enlentecimiento en los ensayos inválidos ocurre en ambos campos visuales, y la única condición es que el movimiento que debe realizar el sujeto desde la señal hasta el objetivo vaya en la dirección opuesta a la de la lesión (Ladavas, 1987).
También los estudios con PET indican que el lóbulo parietal está implicado en procesos atencionales, más en concreto en los cambios del foco atencional. Corbetta, Meizin, Shulman y Petersen (1993) presentaban a los sujetos objetivos en movimiento de tal manera que estos, para responder, debían mover su atención de izquierda a derecha o de derecha a izquierda en alguno de los dos campos visuales. Estos resultados se compararon con otro grupo de sujetos a los que se les presentaban los mismos estímulos, pero el objetivo sobre el que debían responder permanecía fijo en el centro de la pantalla. Las imágenes obtenidas muestran una activación del lóbulo parietal derecho cuando los estímulos aparecen en el campo visual izquierdo, y de ambos lóbulos parietales cuando es en el campo visual derecho donde aparecen los objetivos. Esta falta de paralelismo entre ambos hemisferios puede explicar la evidencia clínica de que los pacientes con daño en el lóbulo parietal derecho sufren un neglect más acusado que los que tienen dañado el lóbulo parietal izquierdo.
Por último, cabe destacar que los efectos que produce el neglect en la visión también pueden ser encontrados cuando al paciente se le pide que informe sobre imágenes que tenga guardadas en memoria (Bisiach, 1992). Si le pedimos que imagine una escena o lugar conocido y que nos diga los objetos que hay en él, sólo prestará atención a los situados en el mismo campo visual en el que está la lesión. Si después le pedimos que cambie la perspectiva desde donde está imaginando la escena por el lado contrario, ahora nos informará de nuevo de los detalles situados en el mismo lado de la lesión, que fueron los que anteriormente no atendió. Esto parece demostrar que el mecanismo que se ocupa del movimiento de la atención cuando se trata de la visión externa, es el mismo, o al menos está en la misma zona cerebral, que el que se ocupa del movimiento atencional en escenas imaginadas provenientes de la memoria.
Movimiento de la atención. Para que el objetivo pueda ser procesado de forma adecuada el sistema atencional debe alinearse con éste, por lo que ha de moverse hasta situarse en la nueva posición. Parecen existir dos formas de cambiar la atención de lugar: por un lado, cuando movemos los ojos de una localización a otra, los sistemas atencionales son arrastrados por la mirada; pero también puede haber movimiento atencional sin que haya movimiento ocular (Posner, 1978). En la tarea atencional expuesta anteriormente, los sujetos deben mantener, durante todo el ensayo, la mirada fija en el punto de fijación, lo que puede confirmarse registrando los movimientos oculares del sujeto. El hecho de que en los ensayos válidos la respuesta sea más rápida que en los neutrales, indica que ha debido existir un movimiento atencional hacia el lugar señalado que ha anticipado la aparición del objetivo.
Parece que los sistemas cerebrales encargados de los movimientos oculares están íntimamente relacionados con los movimientos encubiertos de la atención. Tanto animales como pacientes que tienen dañados los colículos superiores del cerebro medio presentan problemas para realizar voluntariamente movimientos oculares, sobre todo en la dirección vertical (Rafal, 1998). En humanos, los colículos superiores, se ven afectados por una enfermedad llamada parálisis supranuclear progresiva. Si estos pacientes realizan la tarea atencional, se encuentra que, mientras en la dirección horizontal, no existen problemas para que aparezca el efecto facilitatorio de los ensayos válidos, cuando la atención debe moverse verticalmente, el efecto de validez aparece muy tarde. Esto parece indicar que el problema reside en la lentitud con que estos pacientes mueven la atención desde la posición actual hasta la posición objetivo. También se ha hallado que pacientes afectados por parálisis supranuclear progresiva pierden el efecto de inhibición de retorno, lo que apoya la hipótesis de que este tipo de inhibición está relacionado con el mecanismo responsable de los movimientos oculares. Todos estos hallazgos apuntan a una relación entre los colículos superiores y los mecanismos encargados de los movimientos oculares y atencionales.
Operación de enganche. Una vez que la atención se ha situado en la posición en la que se encuentra el objetivo, la siguiente operación será facilitar el procesamiento de este estímulo por sistemas de procesamiento superior. A este mecanismo se le ha identificado con las propiedades facilitatorias e inhibitorias del foco atencional, por lo que los estudios anatómicos se han centrado en las áreas talámicas, implicadas en su control (Crick, 1992). Aunque en la siguiente sección nos ocuparemos ampliamente de los estudios realizados sobre las propiedades del foco atencional, aquí haremos un breve repaso sobre los datos anatómicos y las evidencias empíricas para la existencia de esta operación.
Tres tipos de datos neurofisiológicos relacionan áreas del tálamo (en concreto el pulvinar) con las operaciones de facilitación e inhibición que realiza la atención para la selección de objetivos. Por un lado, se ha comprobado en monos que existen células en estas zonas que aumentan su actividad cuando se realizan operaciones en las que se requieren movimientos encubiertos de la atención. Además, ésta se ve perjudicada cuando se inyecta alguna sustancia que bloquee la actividad de estas células, enlenteciéndose la respuesta a objetivos situados en el lado contralateral de la lesión.
En estudios con humanos que padecen lesiones en el núcleo pulvinar también se han encontrado déficits cuando realizan la tarea atencional referida anteriormente (Rafal & Posner, 1987). En concreto, se produce un aumento en el tiempo de reacción en ensayos válidos cuyo objetivo está situado en el lado opuesto a la lesión. Una vez descartadas explicaciones por problemas de tipo oftalmológico, este enlentecimiento en la respuesta se ha atribuido a un fallo en la operación de facilitar la localización donde aparecerá el objetivo.
Por último, también estudios realizados con PET han corroborado el papel atribuido al núcleo pulvinar. Esta zona se activaba cuando los sujetos debían responder sobre un objetivo que estaba rodeado de distractores, mientras que no lo hacía si este estímulo se encontraba aislado. La actividad en las áreas visuales no parecía sufrir cambio alguno entre un grupo y otro. Así, parece que de nuevo esta zona se relaciona con las operaciones necesarias para realizar la selección entre objetivo y distractores.
Sistema Atencional Anterior
La función asignada a este sistema es detectar y hacer consciente el objeto estimular que ha sido trasmitido por la red posterior del mecanismo atencional. La detección de un estímulo incluye el reconocimiento de su identidad y la realización de las instrucciones u objetivos a llevar a cabo con el mismo (p.e. iniciar la secuencia de operaciones necesarias para presionar o no una tecla dependiendo de si es la palabra objetivo que se buscaba dentro de una lista). Debido a estas propiedades, a este sistema también se le ha llamado red ejecutiva (Posner & Raichle, 1994). La idea de la existencia de un mecanismo de este tipo se ha basado principalmente en la evidencia de la habilidad humana en reorganizar los procesos de pensamiento de acuerdo con las instrucciones o metas y en la incapacidad que muestran pacientes con daño cerebral en la zona del lóbulo frontal para realizar conductas coherentes con un objetivo.
El giro cingulado anterior y la corteza dorsolateral prefrontal (Funahashi, 2001) son las zonas cerebrales en las que parece residir la mayor parte del sistema atencional anterior. Por un lado, tienen conexiones anatómicas con áreas necesarias para realizar su función de control (desde las zonas perceptuales visual y auditiva, hasta áreas frontales y prefrontales, relacionadas con el mantenimiento activo de representaciones de eventos pasados, working memory (Fuster, 2000). Por otro lado, estudios con PET han revelado una clara relación entre la activación de estas áreas y tareas en las que se requiere detección y selección de estímulos y de respuestas (Colmenero, Catena & Fuentes, 2001). La activación del sistema atencional anterior aparece cuando se requiere la detección de estímulos visuales que deben ser discriminados a partir de su forma, color, movimiento o significado (Corbetta, Meizin, Dobmeyer, Shulman & Petersen, 1990); cuando aumenta el número de elementos a seleccionar y cuando hay poca práctica; y en los bloques conflictivos del la tarea Stroop (Pardo, Pardo, Janer & Raichle, 1990; Botvinick, Braver, Barch, Carter & Cohen, 2001).
Sistema Atencional de Vigilancia
El tercer sistema que forma el mecanismo atencional es el encargado de mantener el estado de alerta necesario cuando se requiere que el sujeto responda a estímulos de aparición infrecuente. Los estudios se han realizado con tareas de larga duración con objetivos muy separados en el tiempo o con tareas que utilizan señales de aviso (Parasuraman & Davies, 1984). Con estas tareas se ha comprobado que el organismo sufre una serie de cambios que ayudan en la detección de la señal. Entre los cambios orgánicos destaca el enlentecimiento del ritmo cardíaco, mientras que en el cerebro se reduce la actividad eléctrica global, a la vez que aumenta el flujo de sangre en áreas como los lóbulos frontal y parietal derechos. Estos cambios provocan una mayor velocidad en la detección de los estímulos objetivo, pero también aumenta el número de errores y de respuestas anticipatorias. Este intercambio entre velocidad y precisión sugiere que la alerta no mejora la calidad del procesamiento del estímulo, sino la velocidad de las acciones (Posner, 1978).
El circuito que parece estar implicado en el mantenimiento del estado de alerta está formado por neuronas norepinefrinérgicas que unen el locus couruleus con áreas del lóbulo frontal principalmente derecho (Posner & Petersen, 1990). La implicación de la norepinefrina se ha demostrado con estudios en los que su acción se ha bloqueado, administrando clonidina o guanfancina, y originándose una disminución en la capacidad de los sujetos para mantener el estado de alerta. La implicación del lóbulo frontal derecho ha venido avalada principalmente por estudios con PET, en los que el flujo sanguíneo aumenta en estas áreas cuando el sujeto está en estado de alerta, y estudios de casos clínicos, que muestran deterioro en este tipo de conductas cuando hay lesiones del lóbulo frontal (Posner & Petersen, 1990).
Un aspecto importante en el estudio del sistema de vigilancia es su relación con el sistema atencional anterior. Cuando los sujetos están en estado de alerta, al mismo tiempo que aumenta el flujo sanguíneo en los lóbulos frontales, disminuye en áreas de este segundo sistema, en concreto en el cíngulo anterior (Cohen & cols., 1988). La explicación que se ha dado a este hallazgo es la siguiente: si el sistema necesita responder rápidamente a un objetivo que aparecerá en cualquier momento, el procesamiento de otros estímulos no relacionados con la tarea en sí, sólo puede producir interferencia y retraso en la detección de éste, por lo que una interrupción momentánea del sistema atencional anterior parecería beneficiar la detección. La sensación a la que se ha asociado esta circunstancia es la de tener “la mente en blanco”, intentando evitar que cualquier estímulo o idea interfiera en la detección de la señal.
Atención espacial
La posición de los estímulos en nuestro espacio visual es una propiedad que parece tener un lugar preponderante en el procesamiento de éstos en el cerebro. Muchos experimentos apoyan esta afirmación (Von Wright, 1968; Woods, 1984). El resultado más frecuente muestra cómo la posición espacial de un objetivo es un indicador más efectivo que cualquier otro (color, forma, significado, etc.) La velocidad de procesamiento y la detección de un objetivo mejora considerablemente si anteriormente a su aparición, una señal indica el lugar donde va a aparecer. Este tipo de resultados llevó a suponer que la atención trabaja seleccionando ubicaciones espaciales, facilitando el procesamiento de los estímulos situados en estas. LaBerge (1983) presentaba a los sujetos palabras de cinco letras. En una condición debían categorizar la letra central y en otra la palabra completa. Ocasionalmente aparecía un objetivo en una de las cinco posiciones y los sujetos debían responder lo más rápidamente que pudiesen sobre su aparición. Los resultados mostraron una función en V para los ensayos en los que había que atender a la letra central, mientras que no se encontraron diferencias en los ensayos en los que la categorización debía hacerse sobre toda la palabra. LaBerge sugirió que un “foco atencional” era enfocado en la posición central y facilitaba el procesamiento de cualquier estímulo que apareciera en esa posición.
La metáfora de foco de luz para explicar el funcionamiento del mecanismo atencional ha sido ampliamente utilizada para dirigir la investigación en este campo. Por esto nosotros la utilizaremos como guía en la exposición de la literatura más relevante sobre el tema. Sin embargo, no todos los investigadores están de acuerdo con este supuesto, y sugieren la posibilidad de que la atención también pudiera dirigirse a objetos perceptuales. Fue Duncan (1984) el primero que planteó explícitamente la distinción entre estos dos tipos de teorías, ofreciendo algunas de las primeras evidencias empíricas en favor de estas últimas. Actualmente hay una creencia generalizada de que la selección atencional actúa dependiendo del tipo de tarea a realizar sobre el mapa espacial o sobre la representación del objeto (p.e. Allport, 1989; Humphreys & Riddoch, 1994). Incluso se ha apuntado la idea de que ambos procesamientos puedan ser llevados a cabo simultáneamente, en paralelo (p.e. Styles & Allport, 1986), reflejando los datos encontrados en estudios neurofisiológicos en los que se ha demostrado la existencia de dos vías paralelas que parten del área V1 (Ungerleider & Mishkin, 1982; Posner & Raichle, 1994). Una de ellas proyectaría al lóbulo temporal inferior, y parece la encargada del procesamiento del qué (forma, color, orientación, textura, etc.), y otra lo haría hacia el lóbulo parietal posterior, analizando el dónde, la ubicación espacial del estímulo.
Tamaño del foco atencional
La metáfora de foco atencional llevó a los investigadores a estudiar los rasgos de la atención selectiva que son compartidos entre estos dos diferentes conceptos. Características como la forma, la divisibilidad, el tamaño, son importantes sólo desde una postura radical en lo referente a la utilidad de la metáfora. Sin embargo, seguir este patrón de ideas nos puede servir para introducir los estudios y datos que han surgido en la psicología cognitiva a partir de esta metáfora. Comenzaremos presentando las principales aproximaciones al estudio sobre el tamaño o la forma del foco atencional.
Las primeras investigaciones tenían como meta descubrir el tamaño del foco. Por ejemplo, Eriksen & Eriksen (1974) pidieron a los sujetos que se aprendieran de memoria dos conjuntos distintos de letras. A continuación se presentaban en el centro de la pantalla letras, de una en una, y los sujetos debían responder a qué conjunto pertenecían. A los lados de estas letras objetivo iban apareciendo otras, distractores, que podían pertenecer al mismo conjunto o al opuesto. Se manipuló la distancia de los distractores al objetivo. Los resultados mostraron un efecto facilitatorio cuando los distractores y los objetivos eran del mismo conjunto e inhibitorio cuando pertenecían a conjuntos opuestos. Pero estos efectos disminuían según iba aumentando la distancia entre los estímulos, desapareciendo casi completamente a partir de un grado de ángulo visual. Los autores concluyeron que estos datos apoyaban la idea de un foco atencional, en el que los estímulos que caen dentro de él son procesados a niveles superiores, pudiendo competir entre ellos por la respuesta, mientras que los que caen fuera del foco no son procesados. Además, el tamaño de éste es de un grado de ángulo visual.
Sin embargo, pronto comenzaron a aparecer resultados que indicaban que este tamaño no era fijo, sino que podía variar dependiendo de la tarea a realizar (Eriksen & Yeh, 1985; Eriksen & James, 1986). De esta manera surgió la idea de una atención, no como foco, sino como lente con zoom. Esta nueva metáfora traía consigo, además de un foco de tamaño variable, la explicitación de que la potencia de procesamiento era uniforme en todo el área focal e inversamente proporcional a la extensión de dicha área. Los primeros estudios que apoyaban estos nuevos supuestos manipularon la extensión del foco antes de la presentación de estímulos con distintos tamaños (Larsen & Bundensen, 1978; LaBerge, 1983; Cave & Kosslyn, 1989). Por ejemplo, Cave y Kosslyn (1989) pedían a los sujetos que respondieran si un estímulo que aparecía en pantalla era un rectángulo o un cuadrado. Manipularon la diferencia entre el tamaño que los sujetos esperaban tuviera el estímulo y su verdadero tamaño. Los resultados mostraron un incremento en el tiempo de reacción según aumentaba la diferencia de tamaños entre el esperado y el real. Concluyeron que parecía existir un mecanismo que adaptaba el foco atencional al tamaño esperado del estímulo, produciéndose un retardo en la respuesta si éste debía ser cambiado posteriormente.
Otros datos apoyaban la idea de un sistema con limitaciones en la cantidad de información que puede ser procesada en un momento dado (Eriksen & James, 1986; Egeth, 1977; Castiello & Umiltà, 1990; Shulman & Wilson, 1987). La idea, como ya hemos comentado, es que cuando el foco atencional tiene un tamaño reducido, la información que cae dentro de él es fuertemente procesada, mientras que si el rango es más amplio, se emplean menos recursos en cada una de las localizaciones u objetos estimulares del área abarcada. Uno de los experimentos más significativos en apoyo de esta idea lo realizaron Shulman y Wilson (1987). Ellos presentaban a los sujetos letras grandes hechas a partir de muchas pequeñas (al igual que los estímulos usados por Navon, 1977). En unos ensayos pedían a los sujetos que identificaran las letras grandes y en otros las pequeñas. Justo después de cada ensayo, tenían también que responder sobre un enrejado sinuosidal, compuesto por líneas gruesas (baja frecuencia espacial) o por líneas finas (alta frecuencia espacial). Su predicción era que si la atención realmente funciona como un foco que adapta su tamaño y su resolución espacial a las necesidades del momento, el cambio entre dos tareas que requieren distinta resolución espacial será más lento, ya que se necesitará un tiempo de adaptación. Los resultados confirmaron la hipótesis, encontrándose respuestas más lentas en los ensayos en los que la resolución espacial que se requería para las dos tareas era diferente.
Uno de los últimos aportes en esta línea de investigación ha sido poner en duda el supuesto de que el tratamiento de la información que cae dentro del foco atencional es totalmente uniforme en él. Muchas investigaciones han encontrado datos que respaldan la idea de un sistema que tiene un pico atencional en el centro del área focal, con una disminución continua según nos alejamos del centro (p.e. Shulman, Wilson & James, 1985; Downing & Pinker, 1985; LaBerge & Brown, 1989; Eriksen & James, 1986; Henderson & Macquistan, 1993). En estos modelos de gradiente también existe una limitación de recursos, integrada dentro de una estructura en la que la cantidad de recursos destinados a un estímulo dado depende de la distancia entre su localización espacial y el centro del área focal. Por lo tanto, el modelo de gradiente predice que la interferencia entre un distractor y un estímulo objetivo disminuye en función de la distancia entre ellos (LaBerge, 1995; Johnston & Dark, 1986). Recientemente, la metáfora de gradiente ha ido cambiando a causa de resultados en los que se demuestra que es la carga informativa, y no la distancia espacial, el factor crítico en el declive de la atención (Lavie, 1995). También se ha sugerido que los datos que apoyan los modelos de gradiente pueden ser explicados si asumimos que los sujetos son imprecisos al situar el foco atencional en un lugar exacto del espacio. Si la localización precisa del foco varía de un ensayo a otro, la media de los resultados en todo el experimento simularía la existencia de una atención con gradiente.
Forma del foco atencional
En una rigurosa interpretación de la metáfora de atención como foco de luz o lente zoom, la forma de ésta debería ser redonda u ovalada. Sin embargo, los objetos perceptuales rara vez se presentan de esa forma, bien al contrario suelen ser figuras complejas, a menudo solapadas con otros estímulos distractores. Así, para que el estímulo objetivo pueda ser seleccionado, el foco atencional debería, en principio, poder adaptarse a cualquier forma. Algunos modelos sugieren que el área seleccionada puede efectivamente tomar cualquier forma (Logan, 1996), sin embargo esta posibilidad apenas ha sido explorada experimentalmente.
Uno de los primeros estudios fue el realizado por Podgorny & Shepard (1983). Ellos presentaban a los sujetos una matriz de celdas de 3x3. De ellas, cuatro o cinco eran ensombrecidas. A continuación aparecía un punto y los sujetos debían responder si estaba en el área ensombrecida o no. Los resultados mostraron que los sujetos respondían más rápidamente cuando el área ensombrecida era más compacta (formas rectangulares o cuadradas), lo que permitió a los investigadores concluir que la atención no podía, o le era más costoso, ajustarse a áreas irregulares. Debido a que estos resultados pueden ser explicados por causas no atencionales, en años posteriores se realizaron experimentos para definir si realmente la forma de la atención era circular. Eriksen y James (1986) pedían a los sujetos que identificaran una letra objetivo incluida, con otros siete distractores, en una formación circular. El objetivo solo podía aparecer en los lugares que con anterioridad eran señalados. En una de las condiciones todos los lugares eran marcados, mientras que en la otra solo la mitad de ellos (siempre situados de forma contigua). En los resultados no aparecieron diferencias significativas entre las dos condiciones, lo que permitió concluir que la atención no puede seleccionar solo la mitad del círculo, sino todo él, por lo que podemos deducir que su forma es ovalada.
Sin embargo, también se encuentra en la literatura evidencia a favor de un foco que puede adaptarse de forma más flexible a las exigencias de la tarea. Joula, Bouwhuis, Cooper y Warner, (1991) presentaban a los sujetos tres anillos concéntricos, de distinto tamaño, con ocho posibles posiciones cada uno en las que podría aparecer el objetivo. En el resto de posiciones se presentaban estímulos distractores. Antes de cada ensayo, uno de los anillos era señalado como el lugar más probable de aparición del objetivo. Se observó un efecto facilitatorio de los ensayos válidos (el objetivo aparecía en el anillo señalado) sobre los inválidos igual para todos los anillos. Esto significa que cuando el sujeto esperaba la aparición del objetivo en el anillo central o en el mayor y este aparecía en uno de menor tamaño, el tiempo de reacción aumentaba significativamente. Si la teoría de que el foco atencional tiene forma circular fuera correcta, el tiempo en responder a un estímulo situado en, por ejemplo, el anillo intermedio debería ser igual, o acaso mayor, que el tiempo en responder a uno situado en el anillo inferior. Por ello, los autores concluyeron que la atención puede tomar cualquier forma, incluida, como en este experimento, la forma de anillo.
Divisibilidad del foco atencional
El debate sobre si la atención visual puede o no atender simultáneamente a dos zonas separadas espacialmente, mientras ignora los estímulos situados entre ellas, ha acompañado a la metáfora de la atención como foco prácticamente desde sus inicios. Los primeros experimentos parecían mostrar que esta división espacial es posible (p.e. Shaw & Shaw, 1977; Shaw, 1978). Sin embargo, pronto aparecieron datos que apuntaban otra posible explicación de este fenómeno (Posner, 1980; Eriksen & Yeh, 1985): no había una autentica división atencional, sino un rápido desplazamiento del foco entre las distintas ubicaciones, esto es, un foco dinámico.
Actualmente, sin embargo, parece haber suficientes evidencias que apoyan la idea de la divisibilidad del foco atencional bajo ciertas circunstancias. Por ejemplo, Castiello y Umiltà (1992) demostraron que se puede atender dos zonas separadas espacialmente cuando éstas se encuentran cada una en un campo visual hemisférico distinto. Aunque se criticaron estos resultados arguyendo la posibilidad de que las medias encontradas se produjeron porque los sujetos atendían en cada ensayo a una zona distinta, la presentación de las distribuciones de los tiempos de reacción y las varianzas permiten excluir esta explicación (Castiello & Umiltà, 1992). Esta crítica, sin embargo, es un referente en todos los experimentos cuyos datos parecen apuntar a que ha habido una división atencional por parte de los sujetos.
Otro aspecto que parece influir decisivamente en los datos es la aparición de estímulos distractores situados entre las dos zonas a atender. Kramer y Hahn (1995) propusieron que la causa por la que algunos experimentos no han podido encontrar una división atencional genuina es porque en sus ensayos los estímulos aparecían repentinamente, capturando automáticamente la atención. Kramer y Hahn (véase también Hahn & Kramer, 1998) sugieren que la captura de la atención por parte de estímulos distractores que aparecen repentinamente, puede dificultar o imposibilitar a los sujetos mantener su atención dividida entre las dos zonas objetivo. Para comprobar esta hipótesis presentaron a sus sujetos una tarea en la que debían responder si dos letras eran iguales entre sí o no. En la pantalla aparecían cuatro recuadros alineados. Los de los extremos estaban destinados a los objetivos y los centrales a letras distractoras que los sujetos debían ignorar. En una condición todos los estímulos aparecían de forma repentina dentro de estos recuadros, mientras que en la otra condición las letras se formaban a partir de movimientos de segmentos de figuras que ya estaban en pantalla. Como se predijo, sólo en la condición en la que los estímulos aparecían de forma repentina, los distractores centrales influyeron en la respuesta a los objetivos.
También Bichot, Cave y Pashler (1999) han encontrado, en tareas de búsqueda visual, que los sujetos pueden atender dos zonas espaciales distintas. En definitiva, parece que cuando se dan las condiciones necesarias la atención sí puede ser dividida, pero que esta situación es muy inestable, probablemente por la cantidad de recursos que se requieren, y cualquier demanda, especialmente si es de carácter exógena, devuelve el sistema a un estado unifocal.
Movimiento del foco atencional
Son varios los aspectos que se han estudiado en relación a cómo la atención visual se desplaza a través del espacio. Uno de ellos es su relación con los movimientos oculares. Aunque suele coincidir la orientación de la atención con la de los receptores sensoriales (en particular con la fóvea retiniana), existe amplia evidencia en la literatura a favor de una independencia relativa entre ambos (Posner, 1978; Posner, 1980). Por ejemplo, Posner (1980) comprobó que, sin que se produzca ningún movimiento ocular, los sujetos tienden a detectar más rápidamente objetivos cuya posición de aparición ha sido señalada previamente, que aquellos en los que no aparece señal de aviso alguna. Esta ganancia de tiempo en los ensayos válidos ha sido interpretada como un movimiento encubierto de la atención hacia el lugar donde va a aparecer el objetivo.
A pesar de que la atención pueda moverse de forma independiente al movimiento ocular, existen datos que relacionan ambos. Según Posner y Petersen (1990), estos dos movimientos son llevados a cabo por las mismas estructuras cerebrales, concretamente las que forman parte de la red atencional posterior. Además, parece que en algunos casos la atención ejerce de guía para el movimiento ocular. Por ejemplo, Stelmach, Campsall y Herdman (1997) comprobaron que la amplitud de los movimientos oculares durante la lectura se ajusta para adaptarse a la estructura de la nueva información, lo que parece indicar que la atención se adelanta para preparar este movimiento.
Un segundo aspecto estudiado es la naturaleza discreta o continua del movimiento atencional. Cuando la atención se desplaza entre dos puntos, ¿recorre todos los puntos intermedios procesando, aunque sea sucintamente, los estímulos que hay en el trayecto, o salta de un lugar a otro sin recorrer la zona intermedia? El primer estudio sobre el tema lo realizaron Shulman, Remington y Malean (1979). En cada ensayo presentaban un objetivo, que podía aparecer en ambos lados de la pantalla, sobre el que los sujetos debían responder. Previamente presentaban una señal que indicaba dónde aparecería más probablemente. En algunos ensayos, de forma esporádica, el objetivo en lugar de aparecer a la distancia habitual, lo hacía en una localización que se encontraba entre el punto de fijación y el de la señal. En los ensayos inválidos también el objetivo podía aparecer en esta situación intermedia. Manipularon el tiempo entre la aparición de la señal y la aparición del objetivo. El resultado más interesante fue que la facilitación que se encuentra en los ensayos válidos con respecto a los neutrales e inválidos apareció con intervalos de tiempo más cortos en la posición intermedia que en la de la señal. Esto llevó a los autores a proponer que la atención, cuando se desplaza, pasa por todos los puntos intermedios.
Sin embargo, las conclusiones de este estudio se cuestionaron tanto por problemas metodológicos como por la difícil interpretación de algunos de los resultados colaterales, principalmente los referidos a las diferencias encontradas entre las dos localizaciones de los ensayos inválidos (Eriksen & Murphy, 1987; Yantis, 1988). Esto hizo que aparecieran un buen número de investigaciones tratando de resolver la cuestión. La mayoría utilizaron métodos parecidos a los de la investigación recién comentada. Por ejemplo, Murphy y Eriksen (1987) situaron en la posición intermedia un distractor, razonando que si la atención pasaba por este punto, la ejecución debería verse afectada, mientras que si la atención saltaba hasta la localización del objetivo, la presencia o no de un estímulo distractor no debería afectar. Los resultados apoyaron esta segunda hipótesis, por lo que los autores concluyeron que la atención se mueve de forma discreta, sin pasar por puntos intermedios. Otros experimentos más recientes y con mayor rigor metodológico también apuntan en esta dirección (p.e. Chastain, 1992). La idea que actualmente se tiene es que parece que un cambio atencional espacial se implementaría en el cerebro como un decremento en la actividad de un conjunto de neuronas cuyos campos receptivos cubrieran la zona atendida anteriormente, junto con un incremento en la actividad de un nuevo conjunto de neuronas relacionadas con la nueva localización a atender (Motter, 1994; Schall, Hanes, Thompson & King, 1995).
El tercer aspecto que vamos a tratar con relación a los movimientos atencionales es la velocidad con que se producen. La pregunta es si cuando la atención debe desplazarse de un punto del espacio a otro, el tiempo que emplea es proporcional a la distancia que los separa o es fijo, independientemente de ésta. El primero en explorar esta línea de investigación fue Tsal (1983). Presentaba a sus sujetos dos letras que debían identificar. Podían aparecer en seis posiciones, tres a la derecha del punto de fijación y tres a la izquierda, alineadas horizontalmente. Antes presentaba una señal que indicaba en qué lado aparecería, con más probabilidad, el objetivo. Manipuló la diferencia de tiempo entre la aparición de la señal y la del objetivo (SOA). Como en todos los experimentos de este tipo, para todas las localizaciones el tiempo de reacción aumenta según aumenta el SOA, hasta alcanzar una asíntota. Se supone, que la asíntota en el tiempo de reacción corresponde con el tiempo que tarda el sujeto en desplazar su atención hasta la localización señalada. Los resultados mostraron asíntotas más altas para las distancias mayores. Esto llevó al autor a proponer que la atención se movía con una velocidad constante. Sin embargo, también este experimento fue objeto de críticas metodológicas (ver Yantis, 1988), lo que llevó a otros autores a continuar con la investigación. En años posteriores, y utilizando métodos parecidos, se obtuvieron datos a favor de un foco atencional que se mueve por el espacio a saltos, siendo el tiempo que tarda el mismo para todas las distancias (Remington & Pierce, 1984; Sagi & Julezz, 1985; Eriksen & Webb, 1989; Kwak, Dagenbach & Egeth, 1991). Un resultado idéntico, pero utilizando una aproximación diferente, obtuvieron Sperling y Weichselgartner (1995). Ellos presentaban a sus sujetos, de forma continua, dos tipos diferentes de estímulos: en la fóvea, números, y en la parafóvea, letras. La tarea atencional era detectar el siguiente estímulo que apareciera en la secuencia numérica después de la aparición de una letra objetivo. La tarea motora consistía en apretar un botón en cuanto se detectara la aparición de esta letra objetivo. En anteriores experimentos habían comprobado que mientras la realización de la tarea motora sí alteraba la tarea atencional, esto no ocurría a la inversa. Así, utilizaron los tiempos de reacción de la tarea motora como índice de la detección del objetivo. Manipularon la distancia entre la localización del flujo de letras y la de números. Una vez sustraído el tiempo de detección del objetivo (tarea motora), no se obtuvo efecto de la variable distancia sobre el tiempo de reacción en la tarea atencional. Así, parece que la mayoría de los resultados en la literatura apunta a que los cambios de atención en el espacio se realizan a saltos, sin pasar por las localizaciones intermedias, y empleando el mismo tiempo, independientemente de la longitud del salto.
Un fenómeno que ha sido muy estudiado, y que está relacionado con los movimientos atencionales, es el de la inhibición de retorno. Cuando los investigadores comprobaron que si antes de la aparición de un objetivo, al sujeto se le indicaba su lugar de aparición mediante una señal periférica (un estímulo que aparece en la misma localización), sus tiempos de reacción mejoraban. Pero cuando manipularon el intervalo entre la aparición de los dos estímulos se observó que si era superior a los 300 m los sujetos tardaban más en responder. Este fenómeno fue denominado como inhibición de retorno, ya que se produce un enlentecimiento en el procesamiento de estímulos que están situados en una localización previamente atendida (Posner & Cohen, 1984). El efecto responde a una conducta adaptativa que impide que, en la exploración de una escena, inspeccionemos de forma reiterada un mismo lugar, ya que se supone que en períodos tan cortos de tiempo no debería haber cambios significativos de la información en esa localización. Aunque en la actualidad todavía existen discrepancias con respecto a su naturaleza (por ejemplo, Milliken, Joordens, Merikle & Seiffert, 1998), la mayoría de los autores sostienen que la inhibición de retorno es principalmente atencional y que, además de inhibir localizaciones, el efecto se produciría también sobre objetos, aunque estos estén en movimiento (Tipper, Weaver, Cameron, Brehaut & Bastedo, 1991; Tipper, Weaver & Houghton, 1994).
Procesamiento fuera del foco atencional
El procesamiento de los estímulos que no están situados dentro del foco atencional viene condicionado por el tipo de foco que suponemos que existe (Cave & Bichot, 1999). Si suponemos que tiene una forma de haz de luz (con zoom o sin él), con unos límites definidos, los estímulos que caigan dentro o fuera de su zona de influencia deben recibir un procesamiento cualitativamente diferente. Probablemente sólo los estímulos atendidos pasarán a sistemas de procesamiento superiores, mientras que los demás, o no serán procesados, o este procesamiento se limitará a rasgos determinados. Para que este modelo funcione adecuadamente, el foco deberá poder moverse con rapidez por el espacio, para facilitar los estímulos objetivos situados en distintos lugares. El otro punto de vista es el que considera la atención como un sistema encargado de facilitar el procesamiento de los estímulos situados en un área determinada, pero sin límites definidos. Es el modelo de gradiente. En este caso todos los estímulos son procesados de la misma forma, pero los situados en el centro del área recibirán un plus de facilitación que irá disminuyendo según nos alejamos a la periferia. Este tipo de atención no deberá moverse tan rápidamente por la escena ya que todos los objetivos van a ser procesados, y deja la selección de estos para sistemas superiores de procesamiento.
Los primeros intentos de investigar el tipo de procesamiento que reciben los estímulos situados en la periferia no lograron discriminar entre estas dos posibles explicaciones (p.e. LaBerge, 1983; Downing & Pinker, 1985). Sin embargo, los estudios dirigidos a contrastar ambas hipótesis, han aportado evidencia a favor de las dos. Por ejemplo, Dark, Johnston, Myles-Worsley & Farah, (1985) presentaban palabras previas en dos posibles localizaciones (una de ellas señalada como lugar de aparición del objetivo) durante cortos periodos de tiempo. Inmediatamente después aparecía la palabra objetivo, que estaba relacionada o no semánticamente con la previa. Los tiempos de reacción mostraron efecto de facilitación solo en palabras relacionadas si estas aparecían en la localización objetivo. Estos datos llevaron a los investigadores a concluir que las palabras situadas en la periferia no reciben procesamiento alguno, al menos de carácter semántico. Pero también hay evidencia a favor del procesamiento de estos estímulos. Por ejemplo, Eriksen & James (1986) comprobaron que una letra distractora podía interferir en la respuesta al objetivo, aun estando en posiciones periféricas. Además, cuanto más cerca estuvieron los dos estímulos mayor fue la interferencia, lo que apoya la idea de un foco atencional con gradiente.
Aunque todavía no se ha resuelto definitivamente la cuestión, los datos parecen apuntar a un procesamiento diferente entre los estímulos atendidos y no atendidos, y a la necesidad de tener en cuenta el tipo de tarea con que se estudia. En la literatura se puede observar que los datos provenientes de experimentos con tareas semánticas rechazan la idea del procesamiento de estímulos periféricos (p.e. Johnston & Dark, 1986; Kahneman & Chajczyk, 1983), mientras que en tareas no semánticas sí parece existir ese gradiente de procesamiento o facilitación (p.e. LaBerge & Brown, 1989; Henderson & Macquistan, 1993; Handy, Kingstone & Mangun, 1996). Por tanto, se puede asegurar que los estímulos situados en la periferia del foco atencional son procesados, como mínimo, a nivel de rasgos físicos y espaciales, y que su procesamiento semántico es, al menos, débil (Johnston & Dark, 1986; Cave & Bichot, 1999; Catena, Castillo, Fuentes & Milliken, 2005).
Conclusión
En este trabajo hemos presentado dos de las áreas más importantes en la actualidad de la investigación en atención. La primera es la teoría propuesta por Posner. Las tres redes atencionales permiten explicar y solucionar algunos de los problemas que se han encontrado en la explicación científica de los mecanismos atencionales. Un primer aporte es que las limitaciones de capacidad que estaban en la base de las primeras propuestas han pasado de ser una cuestión relacionada con los sistemas de procesamiento de la información, a ser una limitación de la red atencional anterior. Y son las propiedades que surgen de su propio funcionamiento las que producen esas limitaciones conductuales. En segundo lugar destaca la amplia interacción que se propone entre las tres redes atencionales. Ya hemos hecho mención de éstas, subrayando la influencia continua entre la red posterior y la anterior y el efecto de la activación de la red de vigilancia y las dos anteriores.
En tercer lugar está la importancia que Posner y su teoría le han dado a la neurociencia, como un aliado imprescindible de la psicología cognitiva. A partir de sus estudios, muchos científicos se ocuparon y preocuparon en introducir correlatos neurales en sus teorías psicológicas. Esto produjo un enriquecimiento de éstas y un ajuste en algunos conceptos. Aunque el tema es demasiado amplio como para abordarlo aquí, comentar que esta revolución ha llegado a tal extremo que actualmente las investigaciones y las publicaciones más destacadas y citadas en atención son estudios de imágenes mentales. En todos los principales laboratorios del mundo existen aparatos de neuroimagen, y los investigadores en atención se están convirtiendo en expertos neurocientíficos.
Pero estos mismos cambios están produciendo, a nuestro entender, un efecto negativo sobre la teoría de las redes atencionales. Los estudios de neuroimagen buscan principalmente encontrar las causas de la conducta, y, si se analiza detalladamente la teoría de Posner, se encuentra que ésta tiene más rasgos de una teoría descriptiva que de una explicativa, con pocas predicciones que explorar. La falta de utilidad en este campo, pensamos, predice un abandono futuro no muy lejano, que será más brusco en tanto aparezca otra teoría general más integrada con los nuevos datos neuropsicológicos.
También en este artículo hemos revisado los datos referentes a la atención visual selectiva. Como hemos visto, este campo está dominado por la metáfora del foco atencional, y la mayoría de los estudios se ocupan de demostrar propiedades de la atención relacionados con ella, como son tamaño, forma, movimiento, etc. Esta metáfora ha recibido muchas críticas (Fernandez-Duke & Johnson, 1999; Cave & Bichot, 1999), pero ha sido una buena guía de estudio, produciendo una gran cantidad de datos que han permitido caracterizar su funcionamiento. Sin lugar a dudas, el principal problema que existe en el área es la falta de una teoría que unifique todos estos resultados, y que proporcione una explicación válida a las relaciones con los sistemas de procesamiento posteriores. Los autores confían que los anteriormente citados estudios de neuroimagen faciliten ambas integraciones.
1 Las metas a las que aquí nos referimos no son metas conductuales, sino la realización del tipo de procesamiento que sobre ese objeto estimular debe realizarse (p.e. análisis de alguna de sus características, comparación con otro objeto en memoria, etc.)
Referencias
Allport, D. A. (1989). Visual attention. En M. I. Posner (Ed.), Foundations of cognitive science. Cambridge, MA: MIT Press. [ Links ]
Baddeley, A. D. (1986). Working memory. Oxford, England: Clarendon Press. [ Links ]
Bashinski, H. S. & Bacharach, V. R. (1980). Enhancement of perceptual sensitivity as the result of selectively attending to spatial locations. Perception and Psychophysics, 28(3): 241-248. [ Links ]
Bichot, N. P., Cave, K. R., & Pashler, H. (1999). Visual selection mediated by location: Feature- based selection of noncontiguous locations. Perception and Psychophysics, 61, 403-423. [ Links ]
Bisiach, E. (1992). Understanding consciousness: Clues from unilateral neglet and other disorders. En A. D. Milner y M. D. Rugg (Eds.), The Neuropsychology of Consciousness. Academic Press. [ Links ]
Broadbrent, D. E. (1958). Perception and communication. London: Pergamon Press. [ Links ]
Botvinick, M. M.; Braver, T. S.; Barch, D. M.; Carter, C. S. & Cohen, J. D. (2001). Conflict monitoring and cognitive control. Psychological Review, 108(3), 624-652. [ Links ]
Callejas, A., Lupiañez, J. & Tudela, P. (2004). The three attentional networks: On their independence and interactions. Brain and cognition, 54, 225-227. [ Links ]
Cave, K. R. & Kosslyn, S. M. (1989). Varieties of size-specific visual selection. Journal of experimental psychology: General, 118, 148 - 164. [ Links ]
Castiello, U. & Umiltà, C. (1990). Size of the attentional focus and afficiency of processing. Acta Psychologica, 73, 195 - 209. [ Links ]
Castiello, U. & Umiltà, C. (1992). Splitting focal attention. Journal of Experimental Psychology: Human Perception and Performance, 18, 837 - 848. [ Links ]
Castillo, A. & Catena, A. (2003). Propiedades espaciales del macanismo atencional inhibitorio. Psicología desde el Caribe, 12, 173-202 [ Links ]
Cepeda, N. J., Cave, K. R., Bichot, N. P., & Kim, M. S. (1998). Spatial selection via featuredriven inhibition of distractor locations. Perception and Psychophysics, 60, 727-746. [ Links ]
Catena, A., Castillo, A., Fuentes, L, J. & Milliken, B. (2005). Processing of distractors inside and outside the attentional focus in a priming procedure. Visual Cognition. En prensa [ Links ]
Chastain, G. (1992). Analog versus discrete shifts of attention across the visual field. Psychological Research, 54, 175 - 181. [ Links ]
Cherry, E. C. (1953). Some experiments on the recognition of speech with one and two ears. Journal fo the Acoustical Society of America, 25, 975-979. [ Links ]
Colmenero, J. M., Catena, A. & Fuentes, J. L. (2001). Atención visual: Una revisión sobre las redes atencionales del cerebro. Anales de Psicología, 17(1), 45-67. [ Links ]
Corbetta, M., Meizin, F. M., Dobmeyer, S., Shulman, G. L. & Petersen, S. E. (1990). Attentional modulation of neural processing of shape, color and velocity in human. Science, 248, 1556-1559. [ Links ]
Corbetta, M., Meizin, F. M., Shulman, G. L., & Petersen, S. E. (1993). A PET study of visuospatial attention. Journal of Neuroscience, 13(3), 1202-1226. [ Links ]
Cohen, R. M, Semple, W. E., Gross, M., Holcomb, H. J., Dowling, S. M. & Nordahl, T. E., (1988). Functional localization of sustaided attention. Neuropsychiatry, Neuropsychology and Behavioral Neurology, 1, 3-20 [ Links ]
Crick, F. (1992). Function of the thalamic reticular complex: The searchlight hypothesis. En Kosslyn, S. M. y Andersen, R. A. (Ed.), Frontiers in cognitive neuroscience. (pp. 366-372). [ Links ]
Dalrymple-Alford, E. C. & Budayr, B. (1966). Examination of some aspects of the Stroop color-wrod test. Perceptual and Motor Skills, 23, 1211-1214. [ Links ]
Dark, V. J., Johnston, W. A., Myles-Worsley, M. & Farah, M. J. (1985). Levels of selection and capacity limits. Journal or Experimental Psychology: General, 114, 472-497. [ Links ]
DeRenzi, E. (1982). Disorders of space exploration and cognition. New York: Hohn Wiley & Sons. [ Links ]
Downing, C. J. & Pinker, S. (1985). The spatial structure of visual attention. En M.I. Posner y O.S.M. Marin (Eds), Attention and performance XI: Mechanisms of attention (pp. 171-187). Hillsdale. [ Links ]
Duncan, J. (1984). Selective attention and the organization of visual information. Journal of Experimental Psychology: General, 4, 501-517. [ Links ]
Egeth, H. (1977). Attention and preattention. En G. H. Bower (Ed), The psychology of learning and motivation, 11, pp. 277-320. New York: Acaddemic Press. [ Links ]
Eriksen, B. A. & Eriksen, C. W. (1974). Effects of noise letters upon the identification of a target in a non-search tast. Perception and Psychophysics, 16, 143-149. [ Links ]
Eriksen, B. A. & Murphy, T. D. (1987). Movement of attentional focus across the visual field: A critical look al the evidence. Perception and Psychophysics, 42, 299-305. [ Links ]
Eriksen, B. A. & St. James, J. D. (1986). Visual attention within and around the field of focal attention: A zoom lens model. Perception and Psychophysics, 40, 225-240. [ Links ]
Eriksen, B. A. & Webb, J. M. (1989). Shifting of attentional focus withing and about a visual display. Perception and Psychophysics, 45, 175-183. [ Links ]
Eriksen, B. A. & Yeh, Y. Y. (1985). Allocation of attention in the visual field. Journal of Experimental Psychology: Human Perception and Performance, 11, 583 - 597. [ Links ]
Fan, J., McCandliss, D., Sommer, T., Raz, A. & Posner, M, I. (2002). Testing the Efficiency and Independence of Attentional Networks. Journal of cognitive science, 14 (3), 340- 347. [ Links ]
Fernandez-Duque, D. & Johnson, M. (1999). Attention metaphors: how metaphors guide the cognitive psychology of attention. Cognitive Science, 23, 83-116. [ Links ]
Funahashi, S. (2001). Neuronal mechanisms of executive control by the prefrontal cortex. Neuroscience Research. 2(39), 147-165. [ Links ]
Fuster, J. M. (2000). Cortical dynamics of memory. International Journal of Psychophysiology, 2-3(35), 155-164. [ Links ]
Johnston, J. A. & Darl, V. J. (1986). Selective attention. Annual Review of Psychology, 37, 43-75. [ Links ]
Joula, J. F., Bouwhuis, E. E., Cooper, C. & Warner, B. (1991). Control of attention around the fovea. Journal of Experimental Psychology: Human Perception and Performance, 17, 125 -141. [ Links ]
Hahn, S. & Kramer, A. F. (1998). Further evidence of division of attention over noncontiguous visual field. Visual Cognition, 5, 217-256 [ Links ]
Handy, T. C., Kingstone, A. & Mangun, G. R. (1996). Spatial distribution of visual attention: Perceptual sensitivity and response latency. Perception and Psychophysics, 58, 613-627. [ Links ]
Henderson, J. M. & MacQuistan, A. D. (1993). The spatial distribution of attention following an exogeneous cue. Perception and Psychophysics, 53, 221 - 230. [ Links ]
Holender, D. (1986). Semantic activation without conscious identification in dichotic listening, parafoveal vision, and visual masking: A survey and appraisal. Behavioral and Brain Sciences, 9(1), 1-66. [ Links ]
Hougton, G. & Tipper, S. P. (1994). A dinamic model of selective attention. En D. Dagenbach y T. Carr (Eds.), Inhibitory mechanisms in attention, memory and language, Orlando, FL: Academic Press. [ Links ]
Humphreys, G. W. & Riddoch, M. J. (1994). Attention to within-object and between-object spatial representations: Multiple sites for visual selection. Cognitive Neuropsychology, 11(2), 207-241 [ Links ]
James, W. (1890). The principles of psychology. New York: Dover Publication. [ Links ]
Johnston, W. A., & Dark, V. J. (1986). Selective attention. Annual review of psychology, 37, 43-75. [ Links ]
Kahneman, D., (1973). Attention and effort. Englewoods Cliffs, N.J.: Prentice-Hall Inc. [ Links ]
Kahneman, D. & Chajczyk, D. (1983). Tests of automaticity of reading: Dilution of Stroop effects by color-irrelevant stimuli. Journal of Experimental Psychology: Human Perception and Performance, 9, 497-509. [ Links ]
Kramer, A. F. & Hahn, S. (1995). Splitting the beam: Division of attention over non-contiguous locations of the visual field. Psychological Science, 6, 381-386 [ Links ]
Kwak, H. W., Dagenbach, D. & Egeth, H. (1991). Futher evidence for a time-independent shift of the locus of attention. Perception & Psychophysics, 49, 473-480. [ Links ]
LaBerge, D. (1983). Spatial extent of attention to letters and words. Journal of Experimental Psychology: Human Perception and Performance, 9, 371-379. [ Links ]
LaBerge, D. (1995). Computational and anatomical models of selective attention in object identification. En Gazzaniga, Michael S. (Ed). The cognitive neurosciences. (pp. 649-663). [ Links ]
LaBerge, D. & Brown, V. (1989). Theory of attentional operations in shape identification. Psychological Review, 96, 101-124. [ Links ]
Ladavas, E. (1987). Influence of handedness on spatial compatibility effects with perpendicular arrangement of stimuli and responses. Acta Psychologica. 64(1): 13-23. [ Links ]
Larsen, A. & Bundesen, C. (1978). Size scaling in visual patern recognition. Journal of experimental psychology: Human, Perception and Performance, 4, 1-20. [ Links ]
Lavie, N. (1995). Perceptual load as a necessary condition for selective attention. Journal of experimental psychology: Human, Perception and Performance, 21(3), 451-468. [ Links ]
Lavie, N. (1995). Perceptual load as a necessary condition for selective attention. Journal of experimental psychology: Human, Perception and Performance, 21(3), 451-468. [ Links ]
Logan, G. D. (1996). The CODE theory of visual attention: An integration of space-based and object-based attention. Psychological Review, 103, 603-649. [ Links ]
Marí-Beffa, P. & Nievas, F. (2002). Negative priming from the non-selected meaning of the homograph. British Journal of Psychology, 93, 47-66 [ Links ]
Milliken, B., Joordens, S., Merikle, P. M. & Seiffert, A. E. (1998). Selective attention: Reevaluation of the implications of negative priming. Psychological Review, 105(2), 203-229. [ Links ]
Motter, B. C. (1994). Neural correlates of feature selective memory and pop-out in extrastriate area V4. Journal of Neuroscience, 14, 2190 - 2199. [ Links ]
Murphy, T. D. & Eriksen, C. W. (1987). Temporal changes in the distribution of attention in the visual field in response to precues. Perception & Psychophysics, 42, 576-586. [ Links ]
Navon, D. (1977). Forest before trees: The precedence of global features in visual perception. Cognitive Psychology, 9(3), 353-383. [ Links ]
Neill, W. T. (1977). Inhibitory and facilitatory processes in attention. Journal of Experimental Psychology: Human Perception and Performance, 3, 444-450. [ Links ]
Norman, D. A. (1968). Toward a theory of memory and attention. Psychological Review, 75, 522-536. [ Links ]
Norman, D. A. & Bobrow, D. G., (1975). On data-limited and resource limited processes. Cognitive Psychology, 7, 44-64. [ Links ]
Parasuraman, R. & Davies, D. R. (1984). Varities of attention. New York: Academic Press. [ Links ]
Pardo, J. V., Pardo, P. J., Janer, K. W. & Raichle, M. E. (1990). The anterior cingulate cortex mediates processing selection in the Stroop attentional conflict paradigm, en Proceedings of the National Academy of Sciences, 87, 256-259. [ Links ]
Podgorny, R. & Shepard, R. N. (1983). Distribution of visual attention over space. Journal of experimental psychology: Human, Perception and Performance, 9, 380-393. [ Links ]
Posner, N. I. (1978). Chronometric explorations of mind. Hillsdale, NJ: Lawrence Erlbaum Associates Inc. [ Links ]
Posner, N. I. (1980). Orienting of attention. Quarterly Journal of Experimental Psychology, 32, 3- 25. [ Links ]
Posner, N. I. & Cohen, Y. (1984). Components of visual orienting. En H. Bouman y D. Bouwhuis (Eds), Attention and Performance X, 531-556. Hillsdale, NJ: Erlbaum. [ Links ]
Posner, N. I., Nissen, M. J. & Ogden, W. C. (1978) Attended and unattended processing modes: The role of set for spatial location. En H.J. Pick y I.J. Saltzman (Eds), Modes of perception (pp. 137-157). Hillsdale, NJ: Erlbaum. [ Links ]
Posner, N. I. & Dehaene, S. (1994). Attentional networks. Trends in Neurosciences, 17, 75-79. [ Links ]
Posner, N. I. & Petersen, S. E. (1990). The attention system of the human brain. Annual review of neurosciences, 13, 25-42. [ Links ]
Posner, N. I. & Raichle, (1994). Images of mind. New York: Scientific American Library. [ Links ]
Posner, N. I., Snyder, C. R. R & Davidson, B. J. (1980). Attention and the detection of signals. Journal of Experimental Psychology: General, 109, 160-174. [ Links ]
Rafal, R. D. (1998). Neglect. En Parasuraman, Raja (Ed), The attentive brain, pp. 489-525. [ Links ]
Rafal, R. D. & Posner, M. I. (1987). Deficits in human visual spatial attention following thalamic lesions. Proceeding of the National Academy of Sciences, 84, 7349-7353. [ Links ]
Remington, R. & Pierce, L. (1984). Moving attention: Evidence for time-invariant shifts of visual selective attention. Perception and Psychophysics, 35, 393 - 399. [ Links ]
Sagi, D. & Julesz, B. (1985). Fast noninertial shifts of attention. Spatial Vision, 1, 141-149. [ Links ]
Schall, J. D., Hanes, D. P., Thompson, K. G. & King, D. J. (1995). Saccade target selection in frontal eye field of macaque: I. Visual and premovement activation. Journal of Neuroscience, 15, 6905-6918. [ Links ]
Shaw, M. L. (1978). A capacity allocation model for reaction time. Journal of Experimental Psychology: Human Perception and Performance, 4, 586-598. [ Links ]
Shaw, M. L., & Shaw, P. (1977). Optimal allocation of cognitive resources to spatial locations. Journal of Experimental Psychology: Human Perception and Performance, 3, 201-211. [ Links ]
Shulman, G. L., Remington, R. W. & McLean, J. P. (1979). Moving attention through physical space. Journal of Experimental Psychology: Human Perception and Performance, 5, 522-526. [ Links ]
Shulman, G. L. & Wilson, J. (1987). Spatial frequency and selective attention to spatial location. Perception, 1(16), 103-111. [ Links ]
Shulman, G. L; Wilson, J. S. & James, B. (1985). Spatial determinants of the distribution of attention. Perception and Psychophysics, 1(37), 59-65. [ Links ]
Sperling, G. (1960). The information available in brief visual presentations. Psychological Monographs, 74, 498-515. [ Links ]
Sperling, G., & Melchner, M. J. (1978). Visual search, visual attention and the attention operanting characteristic. En Requin (Ed.), Attention and performance VII, Hillsdale, NJ: Erlbaum. [ Links ]
Sperling, G. & Weichselgartner, E. (1995). Episodic theory of dynamics of spatial attention. Psychological Review, 102, 503-532. [ Links ]
Stelmach, L. B., Campsall, J. M. & Herdman, C. M. (1997). Attentional and ocular movements. Journal of Experimental Psychology: Human Perception and Performance, 3(23), 823-844. [ Links ]
Styles, E. A. & Allport, D.A. (1986). Perceptual integration of identity, location and colour. Psychological Research, 4(48), 189-200. [ Links ]
Tipper, S. P. (1985). The negative priming effect: Inhibitory effects of ignored primes. Quarterly Journal of Experimental Psychology, 37, 571-590. [ Links ]
Tipper, S. P., Weaver, B., Cameron, S., Brehaut, J. & Bastedo, J. (1991). Inhibitory mechanisms of attention in identification and localization tasks: Time course and disruption. Journal of Experimental Psychology: Learning, Memory and Cognition, 17, 681-692. [ Links ]
Tipper, S. P., Weaver, B. & Houghton, G. (1994). Behavioral goals determine inhibitory machanisms of selective attention. Quarterly Journal of Experimental Psychology, 47, 809-840. [ Links ]
Treisman, A. M. (1964). Selective attention in man. British Medical Bulletin, 20, 12-16. [ Links ]
Tsal, Y. (1983). Movements of attention across the visual field. Journal of Experimental Psychology: Human Perception and Performance, 9, 523-530. [ Links ]
Tudela, P. (1992). Atención, En J. L. Fernández- Trespalacios y P. Tudela (Coords.), Atención y percepción. Vol. 3 (pp. 119-163). En J. Mayor y J. L. Pinillos (Eds.), Tratado de Psicología General. Madrid: Alhambra. [ Links ]
Umilta, C. (1995). Domain-specific forms of neglect. Journal of Clinical and Experimental Neuropsychology, 17(2) 209-219. [ Links ]
Ungerleider, L. G. & Mishkin, M. (1982). Two cortical visual systems, En D. J. Ingle, M. A. Goodale y R. J. W. (Eds.), Analysis of Visual Behavior, Mansfield: MIT Press. [ Links ]
Van der Heijden, A. (1981). Short-term visual information forgetting. London: Routledge and Kegan Paul. [ Links ]
Von Wright, J. M. (1968). Selection in visual immediate memory. Quarterly Journal of Experimental Psychology, 20(1) 62-68. [ Links ]
Yantis, S. (1988). On analog movements of visual attention. Perception and Psychophysics, 43, 203- 206. [ Links ]

