











 Spanish (pdf)
Spanish (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mail Cited by SciELO
Cited by SciELO  Cited by Google
Cited by Google  Similars in
SciELO
Similars in
SciELO  Similars in Google
Similars in Google 
 Permalink
Permalink1. Introducción
El Chocó es considerado uno de los lugares con mayor diversidad biológica del planeta, con un gran endemismo y especies potencialmente útiles. Debido a su ubicación en el extremo norte de América del Sur; se ha denominado zona de transición de la mesoflora americana, ya que comparte especies del centro y del sur del continente (Gentry, 1986; Galeano, Suárez y Balslev, 1998a, 1998b, García, Moreno, Palacios, Mosquera y Robledo, 2002, Klinger 2011).
Algunos estudios indican que la composición y diversidad florística de los bosques pluviales tropicales del Chocó son explicadas en gran medida por el clima, el drenaje, la topografía y los suelos (Galeano, 2002; Asprilla, Mosquera, Valoyes, Cuesta y García, 2003; Torres-Torres, Mena-Mosquera y Álvarez-Dávila, 2016; Palacios, Perea, Bellido, Caicedo y Abadía, 2017); tales resultados parecen apoyar la teoría del nicho, que asume que las especies coexisten como respuesta a adaptaciones especificas del hábitat, lo que determina la estructura vegetal de un sitio en específico.
Sin embargo, otros estudios realizados a escala local han sugerido que la diversidad florística de un bosque se ve influenciada por procesos aleatorios o biológicos como la limitación en dispersión de especies (Duque, Cavelier y Posada, 2003; Valencia et al. 2004; Quinto y Moreno, 2014).
Sin duda alguna, la información obtenida hasta el momento ha sentado un precedente en cuanto al conocimiento de la composición florística de este tipo de ecosistema; sin desconocer que aún existen áreas de esta región en las que estos atributos han sido poco estudiados. Uno de estos sitios es el bosque pluvial de la subcuenca del río Munguidó, el cual carece de información confiable para planificación eficiente del territorio (COCOMACIA, 2000).
La presente investigación se circunscribe al estudio de la cobertura boscosa de la subcuenca del río Munguidó, la cual es considerada como un ecosistema de importancia para la conservación del recurso hídrico presente en la región, con altos niveles freáticos que actúan como corredores de dispersión y albergue de la fauna silvestre; adicionalmente este tipo de bosque proveen el hábitat a invertebrados que son fuente importante de alimento para la fauna acuática y terrestre. De allí que se hace necesario emprender alternativas que aporten a su conservación (COCOMACIA, 2000).
La forma más efectiva de aportar a la conservación de este tipo de ecosistema es el estudio de la composición y diversidad florística de los bosques, porque permite comparar las comunidades vegetales en función de su riqueza y la variabilidad de especies y evidenciar aspectos de su ecología. Esta información es útil para la planificación y toma de decisiones sobre el manejo y la conservación de los recursos naturales presentes en el sitio (Cascante y Estrada, 2001). Por ello resulta fundamental el conocimiento del estado de la vegetación, considerando que estos ecosistemas son objeto de aprovechamiento por parte de la comunidad y a que aún siguen cumpliendo importantes funciones ambientales (Martínez, Torres-Torres y Medina, 2015).
El objetivo del presente estudio es caracterizar la composición florística, estructura y diversidad del bosque pluvial tropical de la subcuenca del río Munguidó, municipio de Quibdó, Chocó, Colombia. Esta información será de utilidad para la estructuración de planes de manejo que permitan la conservación del bosque, se puede decir que la presente investigación es importante porque por medio de esta se identifica el estado de conservación de un ecosistema estratégico para región del Chocó biogeográfico, se verifica la influencia de la pluviosidad en la presencia o no de algunas familias botánicas, se explora un ecosistema poco estudiado en el centro del departamento y se suministra insumo para la adecuado manejo del bosque.
2. Marco teórico
La principal característica del bosque pluvial tropical es el gran número de especies pero con una baja abundancia, los cuales además presentan patrones complejos de distribución espacial entre el suelo y el dosel. Una manera de acercarse al conocimiento del nivel de organización de estos bosques, es mediante la comprensión de su estructura que permite evaluar la respuesta de los árboles individuales, de las especies y las comunidades (Alvis, 2009). Según Cantillo (2012) la estructura vegetal es entendida como la forma que presenta el bosque y la disposición en el espacio de los individuos de una comunidad vegetal, la cual se enfoca básicamente en dos componentes: vertical y horizontal. El componente vertical estudia la altura de las comunidades vegetales y es comúnmente conocido como estrato, el cual se define como la porción de la masa vegetal contenida dentro de un límite de altura determinado, y se clasifican en cuatro categorías: arbóreo, arbustivo, sub-arbustivo y rasante. Por su parte, el componente horizontal permite conocer la distribución de la vegetación y su relación con los factores ambientales a lo largo de un gradiente horizontal. En este sentido, Rangel-CH et al. (2004), mencionan que a escala geográfica la vegetación está influenciada por el efecto de la altitud y la variabilidad climática del lugar, por lo que la variación en parámetros climáticos (temperatura y precipitación), y su relación con las características del relieve y de la topografía ejercen un papel determinante en la distribución actual de la vegetación.
Otros investigadores como Magurran (1988), Moreno (2001) y Villarreal et al. (2004) han propuesto métodos de evaluación respecto a los cambios de la biodiversidad con relación a la estructura del paisaje. Estos autores afirman que para monitorear estos cambios se precisa contar con información de la diversidad biológica (diversidad alfa), la tasa de cambio de la biodiversidad entre distintas comunidades (diversidad beta), para conocer su contribución al nivel regional (diversidad gama), y así planificar el manejo del ecosistema y su conservación. La mayoría de técnicas para evaluar la diversidad de especies y su estructura dan comienzo dentro de las comunidades; basándose por un lado en la cuantificación del número de especies presentes (riqueza específica) y, por otro, en la estructura de la comunidad, es decir, la distribución proporcional del índice valor de importancia (IVI) de cada especie (Valoyes, Ramírez, Klínger y Carabalí, 2012; Quinto y Moreno, 2014; Torres-Torres et al. 2016; Torres-Torres, Mena-Mosquera y Álvarez-Dávila, 2017). Así, los autores recomiendan cuantificar el número de especies (riqueza) y su representatividad (estructura), de tal forma que ambos parámetros sean complementarios en la descripción de la diversidad.
La estructura de la vegetación puede abordarse a través de índices que expresan la ocurrencia de las especies, lo mismo que su importancia ecológica (Krebs, 1989). De esta manera, la caracterización de la vegetación con base en las especies dominantes se puede abordar según la fisionomía o la composición florística. En el primer caso, los análisis se dirigen a la evaluación de la estructura horizontal que caracteriza las especies que presentan los mayores valores en tres parámetros ecológicos principales: la abundancia, la frecuencia y la dominancia, cuya suma relativa genera el IVI (Rangel-CH et al. 2004). Las especies con mayores valores de IVI son también llamadas "especies preponderantes", que se definen como aquellas que muestran una gran utilidad y modelan de alguna forma los bosques que habitan (Cantillo, 2012). Este índice permite realizar una real aproximación al ecosistema y su importancia desde una perspectiva ecológica (Alvis, 2009).
Los parámetros que dan origen al IVI se definen de la siguiente manera: la frecuencia de un individuo, es la probabilidad de encontrar dicho individuo en una unidad muestral particular; la abundancia se refiere al número de individuos de la especie en área definida, también puede ser expresada como la densidad; mientras que la dominancia usualmente se expresa en términos del área basal, la cual consiste en el área de la sección transversal del tronco o fuste del individuo a 1,30 m del suelo. La generación del IVI requiere de la transformación de los valores absolutos de dominancia, abundancia y frecuencia a valores relativos, expresados en porcentajes del total. De manera que la suma total de los valores relativos de cada parámetro debe ser igual a 100 y, por lo tanto, la suma total de los valores de IVI debe ser 300. Así, este valor resulta ser mejor descriptor que cualquiera de los parámetros utilizados individualmente, revelando la importancia ecológica relativa de cada especie en una comunidad vegetal. Sin embargo, Matteucci y Colma (1982) sugieren que el estudio del rendimiento o área basal de los árboles tiene mayor significado ecológico que su frecuencia. En tal sentido, en el presente estudio el conocimiento del IVI de las especies forestales, constituye la herramienta principal para el análisis de procesos ecológicos espaciales que suceden al interior del bosque pluvial tropical de la Sub-cuenca del río Munguidó.
2. Metodología
Área de estudio
El estudio se realizó en la jurisdicción del Consejo Comunitario Local del Tambo, ubicado al margen izquierdo del río Suruco, en la subcuenca del río Munguidó, localizada al margen izquierdo del río Atrato, al frente de la cabecera municipal de Quibdó (5° 40" 53,7' N y 76° 48" 44,4 O). De acuerdo con el sistema de Holdridge, esta área corresponde a la zona de vida de bosque pluvial tropical (bp-T), la cual presenta una temperatura promedio de 26°C, una humedad relativa de 94,6% y una precipitación anual que oscila entre 4000 y 10.000 mm y el sitio de muestreo se encuentra 54 m de altitud COCOMACIA, 2000; DIAR, 1983; Holdridge, 1996). Las actividades económicas predominantes en la zona son cultivos transitorios, los sistemas agroforestales (SAF) y el aprovechamiento de madera de bosques naturales (Municipio de Quibdó, 2005) (Ver Figura 1).
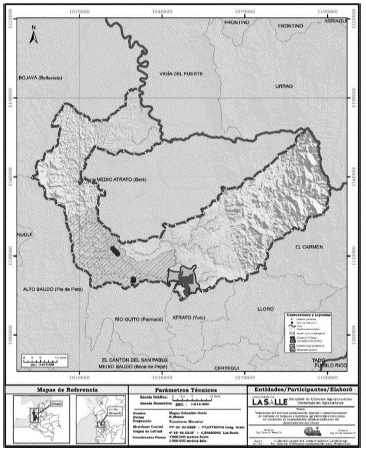
Selección del área de muestreo
El sitio donde se realizó el inventario se ubica a 2,5 km de camino a pie rumbo Sur, detrás del pueblo montaña arriba en el cual se establece el punto de inicio con coordenadas 5° 41' 5,52'' N y 076° 49' 5,74''O, a 107 m de altitud. Establecimiento de unidades de muestreo
Se estableció una parcela temporal de muestreo (PTM) de 1000 x 1000 m (100 ha) en la cual se demarcaron cuatro PTM de 500 x 10 m, empleando la siguiente metodología: el punto de inicio se tomó como punto de partida para trazar una línea base de 1000 m de longitud en dirección norte-sur con la ayuda de una brújula. A medida que se traza la línea base se van colocando estacas cada 50 m hasta los 1000 m. De esta línea base se trazaron cuatro parcelas o unidades de muestreo (L1, L2, L3 y L4) intercaladas en dirección este - oeste utilizando como punto de partida las estacas establecidas a los 50, 350, 650 y 950 m, con longitudes de 500 m de largo por 10 m de ancho (0,5 ha). El área total de muestreo fue de 2 ha, en las cuales se realizaron las mediciones (Ministerio de Medio Ambiente, 2002) (Figura 2).

Fuente: Modificado de Ministerio de Medio Ambiente (2002).
Figura 2 Diseño de parcela para el inventario sistemático.
Muestreo de la composición florística
La composición florística se estudió a través de un inventario sistemático, el cual se encuentra fundamentado en el Decreto 1791/96 del Ministerio de Medio Ambiente (Ministerio de Medio Ambiente, 2002). Se identificaron y registraron las familias, géneros y especies encontradas en el bosque. De igual forma, se midió el diámetro a la altura del pecho (dap) con cinta diamétrica y la altura comercial (hc) y total (ht) con vara graduada, de todos los árboles diámetro a la altura del pecho (dap) ≥ 10 cm, en las unidades de muestreo de 500 x l0 m (0,5 ha) denominadas L1, L2, L3 y L4.
En campo se identificaron todas las morfo especies hasta el máximo nivel taxonómico posible (NN, especie, género, familia botánica), con la colaboración de especialistas en algunos grupos taxonómicos. En el caso de las especies registradas cuya identificación fue desconocida o dudosa, se colectaron tres muestras botánicas (para las estériles) y cinco cuando se disponía de material fértil con su respectivo número de colecta (Galeano, 2002; Torres-Torres et al. 2016). Las muestras botánicas se transportaron hasta el herbario de la Universidad Tecnológica del Chocó "Diego Luis Córdoba", Quibdó, Colombia, donde se confrontaron con el material existente y se identificaron utilizando las claves de Gentry (1993), y Rangel-CH et al. (2004). Posterior a esto, se verificaron la vigencia del nombre científico y la familia en Colombia (Universidad Nacional de Colombia) (http://www.biovirtual.unal.edu.co/ICN/), Missouri Botanical Garden (http://www.tropicos.org) New York Botanical Garden (http://www.nybg.org/), Trópicos (http://www.tropicos.org) y The Internationa/ Plant Names Index (http://www.ipni.org)
Estimación de la composición florística, estructura y diversidad del bosque
Se estimó la abundancia, frecuencia, dominancia dominancia (basado en el área basal), índice de valor de importancia y diversidad, empleando las siguientes ecuaciones:
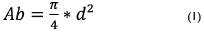
Donde; Ab = Área basal (m2): d2 = diámetro del tronco medido a 1,30 m del suelo al cuadrado (cm)
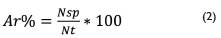
Donde; Ar = abundancia relativa (%), que es la relación porcentual del número de árboles de una especie frente al número total de árboles muestreados; Nsp = número de árboles por especie; Nt: número total de árboles del área muestreada.

Donde; Fr = frecuencia relativa (%), que es la relación porcentual de la frecuencia absoluta de una especie entre la sumatoria total de las frecuencias absolutas de todas las especies; Fasp = frecuencia absoluta de una especie (%); ΣFa = sumatoria de frecuencias absolutas de las especies (%).
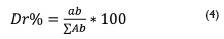
Donde; Dr% = dominancia relativa (%), que la relación porcentual entre el área basal de una especie y la sumatoria total de las áreas basales de las especies; Ab = área basal de cada especie (m2): ΣAb = área basal total en el área de muestreo (m2).
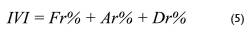
Donde; IVI = índice de valor de importancia; Fr = Frecuencia relativa (%); Ar = Abundancia relativa (%); Dr = dominancia relativa (%).
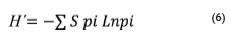
Donde; H' = Diversidad de Shannon-Weaver; S = número de especies; pi = proporción de individuos de la especie i respecto al total de individuos (ni\N); ninúmero de individuos de la especie i dividido entre el número de total de individuos de todas las especies.
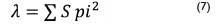
Donde; D = Diversidad de Simpson; S = es el número de especies; pi = proporción de individuos de la especie i respecto al total de individuos.
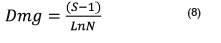
Donde; Dmg = Diversidad de Margalef; S = es el número de especies presentes, N = es el número total de individuos encontrados; Ln = Logaritmo natural.
Adicionalmente, se estimó la distribución diamétrica de especies por clase diamétrica, en donde se organizaron los individuos muestreados con diámetros desde 10 hasta 119,9 cm, en intervalos de 10 cm.
3. Resultados
Composición florística. Se registró una abundancia promedio de 441 individuos/ha, pertenecientes a 71 especies de 62 géneros y 32 familias en un esfuerzo de muestreo total de 2 ha. Las especies de mayor importancia ecológica fueron: Brosimum utile, Virola reiidi y Chrysophyllum argenteum, con un IVI de 22,4; 20,9; 18,2, respectivamente; mientras que las diez primeras especies representan en conjunto un 139,0% del IVI. Los géneros más representativos fueron: Inga, con tres especies, y Clarisia y Brosimum con dos especies cada una. Las familias con mayor representación fueron: Moraceae con 48 individuos/ha de ocho especies y seis géneros; Faboideae con una abundancia de 30 individuos/ha pertenecientes a seis especies y seis géneros y Apocinaceae con 34 individuos/ha de cinco especies y cuatro géneros.
De las 32 familias identificadas, ocho (Moraceae, Faboideae, Apocinaceae, Bombacaceae, Lauraceae, Anonaceae, Mimo-soideaey Meliaseae) representan el 38,4% de la abundancia, lo que indica que existe una alta diversidad de árboles en la zona de estudio (Figura 3).
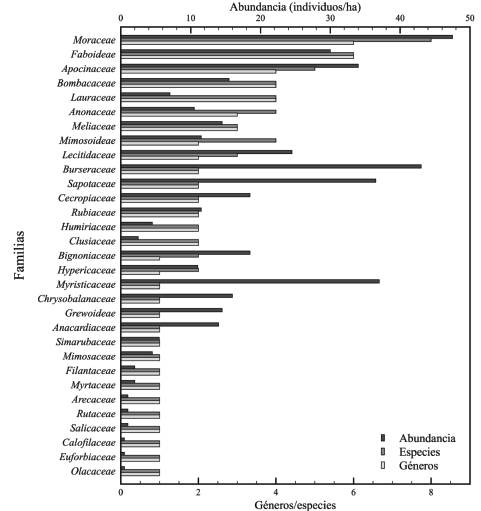
Fuente: Elaboración propia.
Figura 3 Especies y géneros de las familias botánicas más representativas del bosque pluvial tropical de la subcuenca del río Munguidó, municipio de Quibdó, Chocó, Colombia.
Estructura horizontal del bosque
Abundancia. Las especies más abundantes en el bosque de estudio fueron: Virola reiidi, Chrysophyllum argenteum y Protium veneralense con 37 ± 13,9; 36 ± 10,0 y 30 ± 9,1 individuos/ha. Las 10 primeras especies representan el 53,3% de la abundancia en el área muestreada (Figura 2). La abundancia relativa presenta el mismo comportamiento.
Frecuencia. Las especies más frecuentes fueron: Protium veneralense, Pentaclethra macroloba y Chrysophyllum argenteum. De las 71 especies presentes en el área de muestreo, 17 se encontraron en todas las cuatro PTM, nueve en tres, 19 en dos y 24 en una PTM.
Dominancia. Las especies más dominantes fueron: Brosimum utile, Virola reiidi y Chrysophyllum argenteum con 4,6 ± 2,7; 2,6 ± 1,6 y 2,6 ± 0,6 m2/ha, respectivamente. Las 10 primeras especies representan el 60,9% de la dominancia absoluta en el área muestreada (Figura 2).
IVI. Las especies de mayor peso ecológico en el área de estudio son: Brosimum utile, Virola reiidi, Chrysophyllum argenteum con 22,4; 20,9 y 18,3 %. Las 10 primeras especies representan el 138,4% del IVI en el área muestreada (Figura 4).
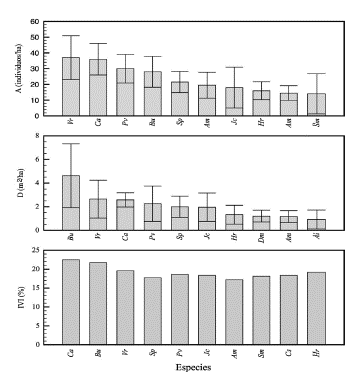
Fuente: Elaboración propia.
Figura 4 Abundancia, dominancia e IVI de especies del bosque pluvial tropical de la subcuenca del río Munguidó, municipio de Quibdó, Chocó, Colombia. A: abundancia; D: dominancia; IVI: índice de valor de importancia. Barras de error corresponden a error estándar Ca = Chrysophyllum argenteum; Bu = Brosimum utile;Vr = Virola reiidi Sp = Schweilera pittieri Pv = Protium veneralense; Jc = Jacaranda copaia; Am = Aspidosperma megalocarpon; Sm = Spondias mombin; Cs = Cecropia spp; Hr = Hirtella racemosa; Dm = Dussia macroprophyllata; Ai = Andira inerms; Am = Apeiba membranacea.
Estructura diamétrica del bosque
Abundancia por clase diamétrica. De acuerdo con el inventario, se encontró que la abundancia presenta una relación inversamente proporcional con el tamaño de la clase diamétrica. Las clases diamétricas menores (10,0-49,9 cm) presentan la mayor abundancia (394 individuos/ha), los cuales se encuentran por debajo del diámetro mínimo de corta (50 cm) y representan el 89,5% de los individuos muestreados. Las clases diamétricas superiores (50,0-119,9 cm) presenta una comparativa menor abundancia: solo 47 individuos/ha y representan el 10,5% de los individuos muestreados (Figura 5).
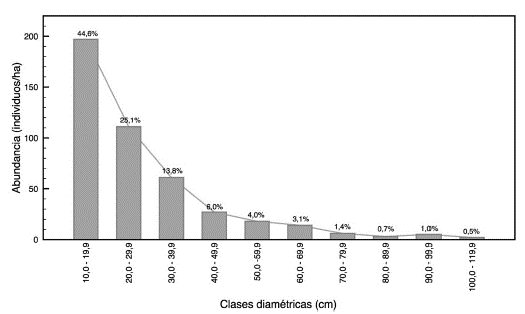
Fuente: Elaboración propia.
Figura 5 Distribución diamétrica del bosque pluvial tropical de la subcuenca del río Munguidó, municipio de Quibdó, Chocó, Colombia.
Área basal por clase diamétrica. Los bosques estudiados presentaron un área basal 34,9 m2/ha, con un promedio de diámetro de 26 cm, siendo 119 cm el diámetro mayor, correspondiente a la especie Virola reiidi de la familia Myristicaceae. Es de notar que existe una alta dominancia de las nueve principales especies, lo cual hizo que influyeran notoriamente en la importancia ecológica de ellas dentro de la comunidad. Estas especies acumularon el 59,3% de la dominancia total de la comunidad.
Diversidad. De acuerdo con los índices de diversidad utilizados, el bosque de estudio presenta una alta diversidad de especies: Equidad de Shannon, Dominancia de Simpson y Diversidad de Margalef de 3,6, 0,03 y 10,3, respectivamente.
4. Discusión
Composición florística. Los resultados obtenidos en este estudio (441 individuos/ha, 71 especies, 62 géneros y 32 familias), suponen una disminución en la composición florística del bosque pluvial tropical en la región del Chocó, si se comparan con el promedio de los resultados registrados por Mosquera, Medina y Martínez, (2012); Palacios et al. (2003); Mena, (2003); Mosquera, Robledo y Asprilla, (2007) (1020 individuos/ha, pertenecientes a 169 especies de 163 géneros y 44 familias). De acuerdo con Andrade (2011), las políticas de estímulo a la ocupación y uso del territorio, la transformación de hábitats y ecosistemas naturales, la ampliación de la frontera agropecuaria, la deforestación, los incendios forestales, el cambio climático y la contaminación, son las causas directas de la pérdida de biodiversidad. Los efectos de la disminución en la composición florística del bosque pluvial tropical, puede ser la pérdida de especies endémicas, hábitat para la fauna asociada y los servicios ecosistémicos de las especies.
La familia Moraceaefue la de mayor importancia ecológica (48 individuos/ha, de ocho especies y seis géneros), no obstante, se destaca una baja presencia de especies de la familia Arecaeae (un individuo/ha, de una especie y un género), contrastando esto con los resultados obtenidos por Gentry (1993); Devia, Cardenas y Cogollo, (1994); Galeano (2002); Mosquera et al. (2012); Valois-Cuesta, Martínez-Ruiz, Rentería y Panesso (2013); Torres-Torres et al. (2016), quienes reportan una alta riqueza de palmas en bosques húmedos tropicales del Chocó biogeográfico. La anterior situación es el resultado de la fuerte presión que ejercen las comunidades del área de estudio a través aprovechamiento de la palma para utilizarla en construcción de viviendas indígenas y como materia prima de artesanías (García et al. 2004). Se hace necesario la implementación de planes de manejo de las especies de la familia Aracaceaeen el área de estudio.
Estructura horizontal del bosque. El IVI del bosque de estudio no se concentra en una o dos familias, ya que cada familia en promedio se encuentra representada por seis individuos/ha, dos especies y dos géneros. Este registro es similar al promedio presentado entre Mosquera et al. (2007) y Mena, (2003) (13 individuos/ha, 2 especies y 2 géneros). Estos resultados son un claro indicador de la estructura y la alta diversidad en el bosques tropicales de la región del Chocó biogeográfico (Valoyes et al. 2012).
Las especies con mayor peso ecológico del bosque estudiado (Brosimum utile, Virola reiidi, Chrysophyllum argenteum, Protium veneralense y Schweilera pittieri), coinciden con las reportadas por Quinto-Mosquera y Álvarez (2010) y Mosquera et al. (2012) en el centro del departamento del Chocó, siendo esto un indicador de la amplia distribución que tienen estos grupos florísticos en esta parte del pacifico colombiano.
Estructura diamétrica. La distribución de las especies por clase diamétrica demostró que el bosque se encuentra en proceso de recuperación o regeneración, hallazgos que resultados similares a los obtenidos por Pardo y Ce-diel (1994), Brown (2000), García et al. (2004), Bermúdez y Blandón (2005), Mosquera et al. (2007). Medina y Martínez (2009) y Rodríguez y Melo (2009), quienes observaron mayor cantidad de individuos en las clases diamétricas inferiores y además sugieren que esta es una de las características de los bosques naturales húmedos tropicales. Estos resultados hacen una invitación a la estructuración de planes de manejo a mediano y largo plazo que favorezca la regeneración natural del bosque y garantice la oferta de productos para el aprovechamiento.
Según Palacios y Ramos (1999) la alta abundancia de individuos de diámetros pequeños o medianos en los bosques del Chocó se debe a la presencia de muchos claros naturales ocasionados por la caída árboles o causados por actividades antrópicas. Por su parte, Pardo y Cediel (1994) consideran que el alto número de individuos con diámetros pequeños se debe a que las especies de este tipo de bosques adoptan estrategias de crecimiento rápido que favorecen la altura antes que el diámetro, además de la extracción selectiva de árboles de diámetros grandes que son económicamente importantes o muestran algún tipo de utilidad Medina y Martínez (2009) afirman que la abundancia de pequeños individuos en las clases diamétricas inferiores indica que el bosque cuenta con potencial para regenerarse y retornar a su estado de evolución anterior. En igual sentido, Mosquera et al. (2007) y Gentry (1993) afirman que los bosques tropicales de la región fitogeográfica del Chocó se caracterizan por una alta densidad de árboles pequeños y medianos (2,5-30 cm de dap).
De acuerdo con Rodríguez y Melo (2009) la estructura del bosque estudiado presenta una línea de tendencia con una distribución diamétrica en forma de "J invertida", lo cual corresponde a una distribución típica y es un indicador de la dinámica del bosque reflejando la característica sucesional de los mismos debido a las perturbaciones constantes y selectivas realizadas por los habitantes de la zona. Los mismos autores afirman que la curva de tendencia de la distribución la población observada, corresponde a una comunidad autoregenerativa, una vez que existe una alta concentración de individuos en las clases menores con reducción acentuada para las clases mayores. Pardo y Cediel (1994) argumentan que dicha tendencia también señala que la comunidad vegetal se encuentra en coherente proceso de desarrollo a etapas de crecimiento y productividad más avanzados, una vez que se confirma la existencia de abundantes individuos jóvenes que irán a suceder árboles cosechados selectivamente o que ya se encuentran posiblemente en la fase senil, pertenecientes a las clases diamétricas superiores. El potencial de regeneración del bosque de estudio, contribuye a mantener su la estructura y diversidad.
Diversidad. El bosque pluvial tropical del departamento del Chocó está catalogado como uno de los de mayor riqueza y diversidad florística en el mundo, en donde en 0,1 ha se han reportado 262 especies de plantas con dap ≥ 2,5 cm (Valoyes et al. 2012). Los valores de diversidad encontrados en este estudio (Equidad de Shannon, Dominancia de Simpson y Diversidad de Margalef de 3,6, 0,03 y 10,3, respectivamente) son comparables a los reportados por Quinto y Moreno (2014) en el bosque pluvial del corregimiento de Salero municipio de Unión Panamericana (Equidad de Shannon, Dominancia de Simpson y Diversidad de Margalef (4,8, 0,01 y 33,2, respectivamente). La diferencia pudo haberse dado por el grado de intervención antrópica e implica que el bosque presenta una alta diversidad de especies, a pesar de la presión antrópica y posible cambio en su composición florística.
5. Conclusiones
El bosque pluvial tropical de la subcuenca del río Munguidó se caracteriza por la presencia de 441 individuos/ha pertenecientes a 71 especies, 62 géneros agrupados en 32 familias botánicas, donde predominan las especies de la familia Moraceae, Faboideae y Apocinaceae y se destaca la baja presencia de especies de la familia Arecaeae. La comparación con otros estudios en este tipo de bosques, evidencian una notoria disminución de la composición florística en estos bosques.
La estructura horizontal del bosque se encuentra definida principalmente por las especies Brosimum utile, Virola reiidi, Chrysophyllum argenteum, Protium veneralense y Schweilera pittieri, las cuales ejercen gran peso ecológico en el bosque y dominan el espacio horizontal. Estas especies son las que más generan resistencia en la perdida de diversidad de especies.
La distribución diamétrica del total de la población es auto regenerativa, dado a que existe una alta concentración de individuos en las clases menores con reducción acentuada para las clases mayores. La anterior situación permite implementar planes de manejo orientados al aprovechamiento selectivo de los individuos de mayor diámetro y altura para abrir claros y contribuir a la regeneración natural.
Según los índices de diversidad utilizados, el bosque de estudio presenta una alta diversidad de especies, a pesar de la presión antrópica y posible cambio en su composición florística. Lo cual es un indicador del potencial auto regenerativo del bosque. Los bosques brindan bienes y servicios ecosistémicos para la humanidad, entre ellos el papel fundamental en la mitigación del cambio climático. Se deben diseñar estrategias de gobernanza que permitan la implementación de una política pública que contribuya a prevenir la pérdida de los ecosistemas boscosos y garantice el manejo sostenible de los mismos.

