











 English (pdf)
English (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mail Cited by SciELO
Cited by SciELO  Cited by Google
Cited by Google  Similars in
SciELO
Similars in
SciELO  Similars in Google
Similars in Google 
 Permalink
Permalink
INTRODUCTION
Vaccinium meridionale, known as the Andean blueberry, agraz or mortiño, belongs to the Ericaceae family and is distributed in northeast South America, predominating in the cold and humid habitats of montane forests between 1,500 and 3,000 m a.s.l. of Colombia, Ecuador, Peru, and Venezuela. In Colombia, wild populations can be found at an altitude up to 3,200 m a.s.l. (Medina et al., 2015; Fischer et al., 2022). The Andean blueberry makes up part of the great natural diversity of plant species in Colombia. This plant has gained importance as a potential promising fruit crop in this country because of nutraceutical and medicinal properties of the fruits, which have a high content of antioxidants (Gaviria et al., 2009; Ligarreto, 2011; Torres et al., 2012; Marín-Echeverri et al., 2018; Garzón et al., 2020). These properties have positioned this plant in different links of the marketing chain since it is widely accepted by growers and pickers and is experiencing growing demand for fresh fruit consumption and processing in the food industry (Quevedo-Rubiano et al., 2021).
This species, also known as the Colombian blueberry (Ligarreto, 2011), has a shrub-like morphology (Ávila et al., 2007) and fruits that are globose berries, green when immature, and dark purple to black when ripe (Magnitskiy and Ligarreto, 2009). Mature fruits are 5 to 10 mm in diameter, with a persistent calyx at the apex (Torres et al., 2012), and contain numerous yellowish-gray to brown seeds. Considering the importance of Vaccinium fruits for human health, consumption of this species has increased in recent years because of market trends (Quevedo-Rubiano et al., 2021). The Andean blueberry is one of the so-called "potential new berries"(Torres et al., 2009).
The antioxidant capacity of berries from the Vaccinium genus, wild or commercially cultivated, depends on the contents of anthocyanins and total phenolic compounds, which, in turn, can be highly influenced by genotype, variations in environmental conditions, fruit maturity stage, and post-harvest storage conditions (Beccaro et al., 2006; Çelik et al., 2008; Vaneková et al., 2020). The importance of the Andean blueberry in the market and fruit nutraceutical and organoleptic characteristics depend on plant origin and territorial biophysical conditions (Ligarreto, 2009; Torres et al., 2009).
These properties mean that production and distribution translate into market potential, both domestic and international, especially since it is marketable internationally (Ligarreto, 2009). Therefore, interest has increased in establishing production systems, to meet internal and external demand. More knowledge on this species is needed to establish a cultivation system under adequate climatic conditions that favor the expression of its genetic potential, while considering all agronomic factors and the adaptability of the taxon to different geographical regions of Colombia (García and Ligarreto, 2014). One of the main limitations for the commercialization of this fruit is the lack of knowledge on optimal practices for crop management and the scarcity of established crops. Since it is a wild species, there are very few records on the relationship between altitude and growth performance (Rada and Cáceres, 2011), with no detailed reports on climatic conditions in Colombia. Therefore, the objective of this study was to evaluate the vegetative growth of Andean blueberry plants at three different altitudes, ranging from 1,885 to 3,298 m a.s.l., in the Department of Cundinamarca. This research sought the least stressful conditions for plant growth along the altitudinal range and practical knowledge on optimal conditions of plant establishment in the field. These data could help to establish optimal cultivation practices for this promising crop, such as selecting the site for crop establishment under favorable climatic conditions.
Cultivation sires and plant material
This study was carried out under field conditions at three sites with different altitudes: i) the township of El Peñón in the municipality of San Francisco - “San Francisco” location at 1,885 m a.s.l. ( 4°57'35.35” N, 74°15'25.28” W), ii) the Faculty of Agricultural Sciences of the Universidad Nacional de Colombia, Bogota - “UNAL” location, at 2,556 m a.s.l. (4°38'9.15” N, 74°5' 25.28” W), and iii) the Cruz Verde páramo - “Paramo” location in the municipality of Choachi at 3,298 m a.s.l. (4°35'33.99” N, 73°59'15.84” W). For the evaluation, 2-year-old plants were used, which were reproduced with in vitro mericlonal propagation. After 10 d of acclimatization in the greenhouse of the Faculty of Agricultural Sciences, Universidad Nacional de Colombia, in Bogota (UNAL location), the plants were planted in plastic pots (35 cm in diameter and 31 cm in height), one plant per pot, to eliminate the influence of the soil factor on plant growth. All pots contained the same soil, with a loamy texture, pH 5.1, organic matter of 9.4%, and an exchangeable acidity of 0.91 cmolc kg-1. In each location, the pots were established in plain-relief sites under free sunlight exposure on December 22 and 26, 2015. At planting in each location, the pots with plants were spaced 30 cm apart to avoid plant shading and incorporated in the soil at a depth of 30 cm. The experiment evaluated vegetative growth for 200 days (d), with measurements taken 18, 70, 114, 163, and 198 d after planting in field.
The experiment was developed with a simple factorial design of repeated measures, with 3 treatments (locations at different altitudes) and 5 samplings, with destructive measurements at the beginning (Tab. 1) and at the end of the trial. In each location, 9 plants were planted in their respective pots. On each measurement date, the following variables were measured: leaf area per plant and plant height according to the methodology described by Lancheros et al. (2012), main stem diameter at the middle-third of the stem (mm), number of primary branches, length of primary branches, and relative chlorophyll content (SPAD) in 5 fully-expanded leaves per plant at the middle-third of the plants. The dry weight of the plants (leaves, stem, roots) was determined at the beginning of the experiment and with the final measurement (198 d) by drying plant organs in oven at 70oC to constant weight.
Table 1. Initial characteristics of plants at establishment of the experiment at three locations with different altitudes.
| Plant height (cm) | Number of primary branches | Number of leaves | Leaf area (cm2) | Shoot (leaves + stems) DW (g) | Root DW (g) | Total plant DW (g) | Relative chlorophyll content (SPAD units) |
|---|---|---|---|---|---|---|---|
| 13.5 | 8 | 62 | 65.73 | 1.147 | 0.696 | 1.843 | 33.7 |
| 17.0 | 2 | 42 | 44.53 | 0.736 | 0.171 | 0.907 | 15.4 |
| 16.5 | 2 | 54 | 57.25 | 0.273 | 0.227 | 0.5 | 22.6 |
| 15.6* | 4* | 52.6* | 55.84* | 0.719* | 0.364* | 1.083* | 23.9* |
*Mean at the beginning of the experiment. DW, dry weight.
Environmental characteristics of the three locations
The environmental conditions relative air humidity and air temperature were recorded using Watch Dog Data Logger® 100 thermo-hygrometers (Spectrum Technologies, Inc., Aurora, IL) placed in each location during the experiment, obtaining data at 30 min intervals. The data were analyzed as an adjusted time series with moving averages to calculate the trend of each climatic variable using the eXtensile Time Series (xts) statistical package in the R software (Ryan and Ulrich, 2012). During the plant growth experiment, the UV solar radiation was measured using an Apogee MU-100 Portable Integrated UV Meter® (ICT International, Armidale, Australia), and the wind speed was measured with a Kestrel 4500 Weather Meter® (Nielsen-Kellerman Company, Boothwyn, PA) at an interval of 10 min. The data were analyzed with descriptive statistics.
The values of air temperature and relative air humidity measured over time for the three locations showed that Paramo (3,298 m a.s.l.) presented the lowest temperature data, with values between 8°C and 10°C on average; UNAL (2,556 m a.s.l.) and San Francisco (1,885 m a.s.l.) presented similar data, with values between 15°C and 22°C. However, in San Francisco, higher temperatures were recorded, where the afternoon hours had average temperatures of 21.0°C. On the other hand, the highest relative humidity was recorded in the Paramo location, with values close to 100%, in contrast to UNAL and San Francisco, with averages of 75.3 and 66.5%, respectively (Fig. 1B).

Figure 1. A) Air temperature and B) Relative air humidity recorded over time at the three altitudes (San Francisco, 1,885 m a.s.l.; UNAL, 2,556 m a.s.l.; Paramo, 3,298 m a.s.l.).
For UV radiation, the highest values were registered in Paramo and UNAL, with means of 81.35 and 82.49 µmol cm-2 s-1, respectively. However, there was a difference in the interquartile range, 41 to 122 µmol cm-2 s-1 for Paramo, and 60 to 99 µmol cm-2 s-1 in UNAL. San Francisco had the lowest mean for UV radiation, with a value of 59.9 µmol cm-2s-1 (Fig. 2A).
Paramo had the lowest wind speed values. In contrast, UNAL and San Francisco had means and interquartile ranges that were very similar, registering a mean of 1.84 and 1.85 m s-1 for San Francisco and UNAL, respectively (Fig. 2B).

Figure 2. Boxplot of UV radiation (A) and wind speed (B) datarecorded at the three altitudes (San Francisco, 1,885 m a.s.l.; UNAL, 2,556 m a.s.l.; Paramo, 3,298 m a.s.l.). The lower base of the box corresponds to Q1, the upper base of the box corresponds to Q3, the line inside the box is the median or Q2 and the point is the arithmetic mean, the upper limit corresponds to Q3+interquartile range (Q3-Q1) *1.5, and the lower limit corresponds to Q1-interquartile range (Q3-Q1) *1.5.
The graphical comparisons were made with variation curves as a function of time. The graphs show the data as the means ± standard deviation; n = 9. The Pearson correlations between the selected variables were calculated, and differences with P≤0.05 were significantly different.
RESULTS AND DISCUSSION
Plant growth variables
The plants grown in UNAL and San Francisco had the highest height and largest leaf area with respect to the plants from Paramo (Fig. 3). The accumulation of DW after 198 d of growth in each location was higher for the plants from UNAL than those from San Francisco, and Paramo had averages of 17.6, 12.9, and 7.8 g DW per plant, respectively. Likewise, the plant height, number of branches, and leaf area had the highest values in plants grown in the UNAL location (Fig. 3).
None of the environments had a significant effect on the length of the primary branches (Fig. 3D). However, the plants in UNAL and Paramo tended to have longer primary branches, while the plants in San Francisco had the shortest primary branches on average (Fig 3D). However, the plants in San Francisco and in UNAL were taller than those grown in Paramo, where the plants had shorter internodes. This indicated that the architecture of these clonal plants was influenced by solar radiation and other climatic factors of the environments (Kawamura and Takeada, 2004). UV-light is a known signal to induce plant morphological responses (Qian et al., 2020). Increased solar radiation at high altitudes commonly reduces internode growth, so plants become shorter and have a “stocky” appearance (Berlia et al., 2013).

Figure 3. Plant height (A), total leaf area per plant (B), number of primary branches (C) and length of primary branches (D) for the Andean blueberry plants grown at three different altitudes (San Francisco, 1,885 m a.s.l.; UNAL, 2,556 m a.s.l.; Paramo, 3,298 m a.s.l.). Data are means (n=9) ± standard deviation.
On the other hand, although the plants from UNAL tended to accumulate a greater average leaf area in relation to the plants from Paramo (Fig. 3B), the UNAL plants had a smaller number of leaves per plant (data not presented). The area of individual leaves was the highest in San Francisco, and the lowest area was in Paramo (data not presented), indicating that, at higher altitudes, there was a reduction in the leaf area of individual leaves. The plants in the Paramo environment had the lowest height (Fig. 3A) and the smallest average stem width (Fig. 4).

Figure 4. Boxplot of the stem width measurements recorded on day 198 of the experiment in the Andean blueberry plants grown at the three altitudes (San Francisco, 1,885 m a.s.l.; UNAL, 2,556 m a.s.l.; Paramo, 3,298 m a.s.l.). The lower base of the box corresponds to Q1, the upper base of the box corresponds to Q3, the line inside the box is the median or Q2 and the point is the arithmetic mean, the upper limit corresponds to the Q3+interquartile range (Q3-Q1) *1.5, and the lower limit corresponds to the Q1-interquartile range (Q3-Q1) *1.5.
Dry weight accumulation in plants
The distribution of DW in plants after 198 d of growth in each of the locations is shown in figure 5. In San Francisco, there was a greater distribution of dry weight towards the roots (34.1%) than in the other locations (23.1-33.5%). In Paramo and UNAL, a greater distribution of dry matter was seen towards the leaves. Plants from San Francisco had a higher percentage of stem dry weight than the other locations (Fig. 5), implying that plants in San Francisco presented more secondary growth than plants from other sites. At the same time, the plants from San Francisco had a lower percentage of dry weight of leaves than the other locations (Fig. 5), which could be attributed to an environmental limitation for emission and growth of leaves at this altitude. An allocation of dry matter towards the stem in the San Francisco plants was, possibly, due to an increase in the efficient use of water (Rodríguez-Castillo and Melgarejo, 2015) under the adverse conditions of San Francisco, contrasting with the other two sites.
The average length of primary branches was higher for the UNAL plants at the end of the experiment, and the UNAL plants tended to produce more branches (Fig. 3C-D). Thus, the UNAL location had an increased number of branches and increased branch length, which explained the greater leaf area per plant (Fig. 3B) and the greater percentage of biomass accumulated in leaves (Fig. 5). These results were similar to those reported by Kawamura and Takeada (2004), who indicated that architecture is influenced by stress and solar radiation, with the influence observed for the average number of branches.
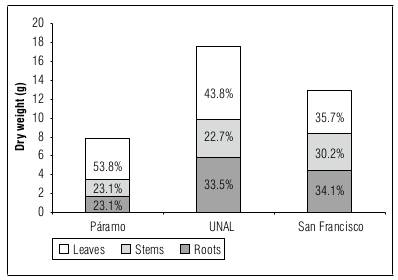
Figure 5. Distribution of dry weight between organs of Andean blueberry plants recorded on day 198 of the experiment at the three altitudes (San Francisco, 1,885 m a.s.l.; UNAL, 2,556 m a.s.l.; Paramo, 3,298 m a.s.l.). Each bar corresponds to the arithmetic mean and shows the percentage distribution over the total.
Chlorophyll contents in leaves
The relative content of chlorophyll in leaves (SPAD) was higher in UNAL than in San Francisco after the 4th sampling on day 114 (Fig. 6). Higher average chlorophyll contents on day 198 were found in the plants from UNAL, with a mean of 50.9 and a standard deviation of 8.5, followed by the plants from San Francisco and Paramo, with means of 43.4 and 36.5 and with deviations of 12.8 and 9.9, respectively. Likewise, after the acclimatization process of the first sampling, the plants had an increase in the anthocyanin contents in leaves (data not present), with greater anthocyanin accumulation in plants in Paramo and San Francisco because of the environmental stimulus. This effect could be attributed to a higher incidence of UV light in the Cruz Verde páramo environment (3,298 m a.s.l.) and the stressful conditions (e.g., elevated air temperatures) in the San Franscisco location (1,885 m a.s.l.). In Vaccinium plants, air temperature and light conditions play an important role in synthesis of phenolic compounds, such as anthocyanins (Karppinen et al., 2016), where plants increase the accumulation of anthocyanins as altitude increases (Zoratti et al., 2015).

Figure 6. Changes in relative leaf chlorophyll content (SPAD) in the Andean blueberry plants grown at different altitudes (San Francisco, 1,885 m a.s.l.; UNAL, 2,556 m a.s.l.; Paramo, 3,298 m a.s.l.). Each sampling date corresponds to the arithmetic mean, the bar in each point corresponds to the standard deviation (n=9).
In general, when comparing plants from the three altitudes, average data of plant height, number of primary branches, leaf area per plant (Fig. 3), plant dry matter (Fig. 5), and relative content of chlorophyll (Fig. 6) were higher in the plants from UNAL. The chlorophyll readings in the leaves of Andean blueberry at the beginning of the experiment (Tab. 1; Fig. 6) were comparable with the relative chlorophyll contents in leaves of deciduous Vaccinium species (Han et al., 2022; Retamal-Saldago et al., 2022). In the UNAL location, the relative chlorophyll contents in leaves tended to increase during the experiment (Fig. 6).
Favorable environmental conditions, such as adequate solar radiation, benefit photosynthetic activity in Vacciniumcorymbosum plants (Petridis et al., 2021). According to our results, San Francisco had less solar radiation and less light energy that might reach the reaction centers, generating a lower photosynthetic activity in the PSII (Rodríguez-Castillo and Melgarejo, 2015). Also, in stressful environments, the decomposition of chlorophyll is possible due to a higher activity of chlorophyllase (Kim et al., 2011). As reported by Kim et al. (2011) for blueberry plants, leaves acclimatized to low solar radiation have a larger area than leaves subjected to higher levels of radiation. This could be the case in the differences in leaf area found between the plants from Paramo and UNAL (Fig. 3B). In the Paramo location, the synthesis of chlorophylls may be affected by low temperatures, as reported by Baizan et al. (2013), while the production of anthocyanins is favored by increasing altitude, which is accompanied by low temperatures (Wang et al., 2014). Additionally, the synthesis of flavonoids is affected by stress conditions since they are antioxidants that protect fatty acids, proteins, and other cell components from oxidative damage (Valledor et al., 2012).
The vegetative growth in the Andean blueberry plants was modified by the environmental factors that exerted continuous pressure on the plants, where light, apparently, was one of the determining factors of growth changes because it affects the carbohydrate synthesis, which would be further reflected in dry matter accumulation (Retamal, 2014). However, photosynthesis is influenced by other climatic factors that affect gas exchange, as indicated by Muraoka and Koizumi (2005). This was observed in the Andean blueberry plants in Paramo, where an average radiation of 81.35 μmol cm-² s-1 was recorded; however, these plants presented the lowest height and smallest leaf area for the three locations. A reduction in leaf area because of increased solar radiation with increasing altitudes has been observed earlier in other species (Derebe et al., 2019; Fischer et al., 2022).
For the distribution of dry weight, the San Francisco plants evidenced dry matter accumulation in the roots, which can be an adaptation attributed to an increase in the efficient use of water under adverse conditions. Also, the elevated soil temperatures in San Francisco, might have benefited root growth. In San Francisco, formation of extensive leaf area would imply increased losses of water during stomatal transpiration. This could be one of reasons for the smaller leaf area in San Francisco than in UNAL. For transpiration, the concentration of water in a stomatal cavity should be greater than the concentration of water vapor in the atmosphere (Rodríguez-Castillo and Melgarejo, 2015); here, San Francisco was different from the other two locations, especially Paramo, where relative humidity values were close to 100% (Fig. 1B).
Correlations between the variables
Significant correlations were found between several morphological variables in the plants (Tab. 2). The stomatal density in the leaves presented a positive correlation with plant height (data not presented), and a positive correlation was found between leaf area and plant height (Tab. 2). Additionally, positive correlations were observed between leaf area and root dry weight, and leaf area and total plant dry weight (Tab. 2). Finally, positive correlations were found between environmental factors, such as UV radiation and air temperature, with total plant dry weight, but relative air humidity negatively correlated with total plant dry weight (Tab. 3).
Table 2 Correlations between response variables in Andean blueberry plants grown at three different altitudes.
| Plant height (cm) | Leaf area (cm2) | Relative chlorophyll content (SPAD) | Root dry weight (g) | Total plant dry weight (g) | |
| Plant height (cm) | Rho=0.63 P=0.0001 | Rho=0.13 P=0.11 | Rho=0.68 P=0.003 | Rho=0.63 P=0.002 | |
| Leaf area (cm2) | Rho=0.63 P=0.0001 | Rho=0.18 P=0.05 | Rho=0.66 P=0.001 | Rho=0.75 P=0.0001 | |
| Relative chlorophyll content (SPAD) | Rho=0.13 P=0.11 | Rho=0.18 P=0.05 | Rho=0.43 P=0.05 | Rho=0.57 P=0.001 | |
| Root dry weight (g) | Rho=0.68 P=0.003 | Rho=0.66 P=0.001 | Rho=0.43 P=0.05 | Rho=0.94 P=0.0001 | |
| Total plant dry weight (g) | Rho=0.63 P=0.002 | Rho=0.75 P=0.0001 | Rho=0.57 P=0.001 | Rho=0.94 P=0.0001 |
Rho is the Pearson correlation coefficient; P is the P-value with a significance of 0.05.
Table 3 Correlations between the morphological variables of Andean blueberry plants grown at different altitudes and environmental factors.
| Plant height (cm) | Plant dry weight (g) | Relative chlorophyll content (SPAD) | |
| Relative air humidity (%) | Rho=0.017 P=0.85 | Rho=-0.64 P=0.002 | Rho=-0.27 P=0.002 |
| Air temperature (°C) | Rho=0.22 P=0.01 | Rho=0.59 P=0.005 | Rho=0.18 P=0.05 |
| UV radiation (µmol cm-2 s-1) | Rho=0.13 P=0.13 | Rho=0.52 P=0.01 | Rho=-0.015 P=0.89 |
| Wind speed (m s-1) | Rho=0.07 P=0.42 | Rho=0.44 P=0.05 | Rho=0.17 P=0.05 |
Rho is the Pearson correlation coefficient; P is the P-value with a significance of 0.05.
The presence of correlations between the variables related to the photosynthetic organs and the environmental factors indicated that the environmental conditions were determinants of the variabilities and morphological and physiological responses of the plants. The fact that branches (ramification) and leaf area are fundamental in the photosynthetic processes of plants indicates that the greater their number is, the greater the carbohydrate synthesis is, favoring the accumulation of dry matter in Andean blueberry plants, depending on the environment.
Plants respond to environmental factors, which can promote or restrict the photosynthetic activity and chlorophyll content needed for optimal growth and development (Rodríguez-Castillo and Melgarejo, 2015). It was evidenced that the plants established at the altitude of Paramo (3,298 m a.s.l.) had less growth than plants from UNAL. This could be explained by the interaction between plants and their respective environments. The metabolic processes of the plants in Paramo diminished, affected by environmental conditions. In contrast, the production of anthocyanins was favored by the increase in altitude, which was accompanied by low temperatures (Wang et al., 2014), as reflected in the reddish tones of the leaves on plants from Paramo.
CONCLUSIONS
Environmental factors affect the morphological responses of the Andean blueberry plants, where plant height, dry matter accumulation, leaf area, and relative chlorophyll content were favored by the UNAL environment at an altitude of 2,556 m a.s.l. On the other hand, San Francisco (1,885 m a.s.l.) had plants that directed their assimilates mainly towards the stem and roots, probably, to avoid losses of water involving the emission and maintenance of new leaves, because of the stressful conditions of this location.
Likewise, although the Andean blueberry plants tended to adapt their growth to the conditions of Paramo (altitudes higher than 3,000 m), in the Cruz Verde páramo, they did not express their physiological potential in terms of growth and, possibly, production.
Changes in growth and fruit production during the reproductive phase of Andean blueberry plants under different environmental conditions need to be evaluated considering the results this study.

